Счет 42 в бухгалтерском учете: торговая наценка
Такое понятие, как торговая наценка, применяется предприятиями, занимающимися розничной коммерческой деятельностью. Именно в целях ведения учета используемых надбавок к цене продаваемого товара, и существует 42 счет. Сегодняшняя тема посвящена определению и значению таких надбавок, и значению 42 позиции для их учета. Мы также рассмотрим типовые бухгалтерские записи по данному счету и разберем один из случаев, который мог бы произойти в жизни, и где применяется данная позиция.
Определение и значение торговой наценки
Как известно, успех коммерческой деятельности во многом определяется применяемой ценовой политикой, а именно гибким подходом к ценообразованию при реализации товаров. Отпускная цена, в свою очередь, является суммой стоимости, которая была уплачена изготовителю либо оптовому поставщику, и надбавки, сумма которая впоследствии должна покрыть понесенные расходы и позволит получить прибыль. Именно от этой добавленной стоимости к покупной цене продукции зависит рентабельность торгового предприятия.
Именно от этой добавленной стоимости к покупной цене продукции зависит рентабельность торгового предприятия.
В различных отраслях экономики размер такой добавленной стоимости определяется по-разному в зависимости от специфики бизнеса. Рассчитать данный показатель, конечно же, можно и используя компьютер. В розничных торговых организациях, например, размер маржинальной прибыли рассчитывается автоматически по каждой реализованной единице товара. Благодаря этому, руководитель службы бухгалтерского учета намного легче определяет конечный финансовый результат. Тем не менее важно понимать, что программное обеспечение с подобными возможностями стоит совсем недешево. Поэтому позволить его себе может далеко не каждая организация. Если речь идет о небольших магазинах либо торговых палатках, то они рассчитывают значение обозначенного показателя вручную.
Таким образом, определить размер получаемой выручки за вычетом себестоимости реализуемой продукции можно по общему товарообороту, по разнообразию реализуемых продуктов, по видам остатков продукции либо по среднему проценту.

Значение обозначенной позиции Плана счетов
42 позиция Плана счетов призвана обобщать сведения о надбавках к стоимости покупки на продаваемую продукцию на предприятиях, торгующих в розницу, которые осуществляют их учет по цене их реализации.
По указанной позиции также ведется учет тех уценок, которые были предоставлены продавцами организациям, занимающимся коммерческой деятельностью в розницу, на случай возможных потерь продукции либо возмещение дополнительных транспортных издержек.
По кредитовой части счета отражается сумма добавленной стоимости принятых к учету товаров.
Размер уценки или накидки по той продукции, которая реализована, отпущена либо списана в результате бракованности товара, его порчи, недостаточности в поставленном объеме и т.д., сторнируется, что оформляется такой бухгалтерской записью, как:
1) Дт 42
Кт 90.
Аналитика по данной позиции должна позволить вести раздельный учет размера скидок и накидок, и разницы в стоимости, относящейся к товарам в организациях розничной коммерческой деятельности, а также к отгруженным товарам.
Стандартные бухгалтерские проводки
Для учета сумм коммерческих уценок и наценок бухгалтерская служба предприятия делает следующие типовые записи:
1) Дт 41
Кт 42 – начисление коммерческой надбавки;
2) Дт 90.2
Кт 42 – списание коммерческой надбавки по проданным товарам;
3) Дт 91.2
Кт 41 – списана сумма превышения уценки над наценкой.
Практический случай
Некое предприятие приобрело партию утюгов в количестве 150 единиц. При этом, общая стоимость покупки составила 329 300 р., сумма НДС – 49 395 р. Размер коммерческой надбавки был определен как 30%. При определении размера торговой наценки и продажной цены товара бухгалтером предприятия были сделаны соответствующие расчеты:
- Размер наценки на всю партию продукции = ((329 300- 49 395))*30%) = 83 972 р.
- Продажная стоимость всей партии = 329 300 – 49395 + 83 972 = 363 877 р.
- Розничная стоимость единицы продукта = 363 877 / 150 = 2 426 р.
При этом были введены следующие проводки:
1) Дт 41
Кт 60 – 279 905 р.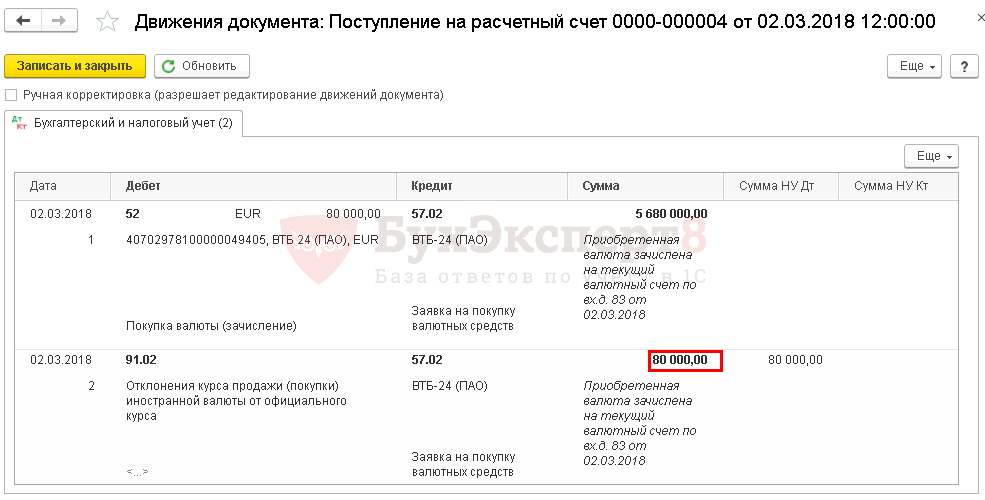 , учет поступившей партии товара;
, учет поступившей партии товара;
2) Дт 19
Кт 60 – 49 395 р., учет суммы НДС;
3) Дт 60
Кт 51 – 329 300 р., перечисление средств за купленные товары;
4) Дт 68
Кт 19 – 49 395 р., НДС к вычету;
5) Дт 41
Кт 42 – 83 972 р., учет торговой наценки.
Заключение
Таким образом, рентабельность деятельности любого торгового предприятия должна обеспечиваться грамотной ценовой политикой. А для того, чтобы руководство предприятия имело четкое представление о ситуации и могла своевременно реагировать на сложившуюся ситуацию, необходимо правильное ведение бухгалтерского учета данного аспекта деятельности.
Счет 42 в бухгалтерском учете — торговая наценка
Общая характеристика счета «Торговая наценка»
Торговая наценка (ТН) — дельта между розничной и закупочной ценой продукции, которая необходима для покрытия издержек и получения прибыли компаниями, осуществляющими розничную торговлю.
Продажная цена формируется из стоимости приобретенных изделий и наценки, которую предприятие определяет самостоятельно. При использовании счета 42 в бухгалтерском учете товары по ценам реализации учитываются по дебету счета 41, а наценка — по кредиту счета 42.
При использовании счета 42 в бухгалтерском учете товары по ценам реализации учитываются по дебету счета 41, а наценка — по кредиту счета 42.
ТН может быть единой для всех видов товаров либо же устанавливаться на каждую номенклатурную позицию (или группу позиций) отдельно.
Рассмотрим общую характеристику счета «Торговая наценка».
Более полную информацию по теме вы можете найти в КонсультантПлюс.Пробный бесплатный доступ к системе на 2 дня.
Учет товаров компания ведет по ценам приобретения или по продажным. Способ учета необходимо закрепить в учетной политике. Во втором случае как раз и необходим отдельный учет ТН, отражающейся проводками по счету 42 бухгалтерского учета.
Многих бухгалтеров интересует вопрос: счет 42 активный или пассивный. Расскажем в следующем разделе.
Счет 42 — активный или пассивный?
Этот счет пассивный, т. е. может иметь только кредитовое сальдо. Более того, дебетовые обороты для него вообще не предусмотрены действующим Планом счетов.
е. может иметь только кредитовое сальдо. Более того, дебетовые обороты для него вообще не предусмотрены действующим Планом счетов.
Счет содержит все сведения о торговых наценках или скидках, учитываемых при расчетах с покупателями. Сальдо по счету показывает величину ТН, относящуюся к непроданным остаткам товаров.
Подпишитесь на рассылку
Аналитический учет по счету 42 должен обеспечивать отдельный учет сумм ТН и скидок.
По экономическому смыслу счет 42 является регулирующим (контрактивным). Самостоятельного значения он не имеет и применяется для уточнения данных, содержащихся на счете 41.
Торговая наценка не отражается в бухгалтерском балансе. Строка 1210, которая показывает запасы организации, содержит сумму без ее учета. Поэтому увидеть и проанализировать ТН можно лишь на счете 42 «Торговая наценка».
Проводки по счету 42
Кредитовые проводки по счету 42 «Торговая наценка» отражают суммы начисленной ТН при оприходовании товаров.
Дебетовые обороты по данному счету не предусмотрены действующими нормативными документами. Поэтому в случае необходимости торговая наценка списывается не прямой записью в дебет этого счета, а сторнированием:
- Дт 41 — Кт 42 — сумма ТН по поступившим товарам;
- Дт 90.2 — Кт 42 (сторно) — ТН по реализованным товарам;
- Дт 44 — Кт 42 (сторно) — списание ТН по использованным для собственных нужд товарам;
- Дт 94 — Кт 42 (сторно) — ТН по товарам, выбывшим в результате порчи или недостачи.
***
Использование счета 42 в бухгалтерском учете нужно для калькуляции торговой наценки. Он применяется на предприятиях, занимающихся розничной торговлей, где учет товаров ведется по продажным ценам. Счет является пассивным, сальдо по нему показывает наценку по непроданным товарам.
***
Больше информации по теме — в рубрике «Бухгалтерский учет».
Розничная торговля
Доброго времени суток.
Подскажите кто чем может!!!
Д41/2 К 41/1 переданы товары в магазин ТОРГ-13;
Д50 К 90 Поступила розничная выручка
…. А дальше не знаю… Как списывается товара со сч. 41/2??? Какими документами это все отражается??? Правильно ли я написала проводки выше??
Буду признательна за любую ПОМОЩЬ!!!
Комментарии
| Розничным торговым организациям дано право учитывать в бухгалтерском учете товары как по покупной, так и по продажной стоимости. Если товар учитывается на счете 41 по покупной стоимости, то его списание ограничивается проводкой: Д 90-2 «Себестоимость продаж» К 41-2 «Товары в розничной торговле». В этом случае валовой доход торговой организации (или доход от продаж, или выручка) определяется как разница между товарооборотом без вычитаемых налогов и себестоимостью проданных товаров. Этот показатель автоматически формируется на счете 90 «Продажи» и складывается из товарооборота, за минусом НДС, акцизов и покупной стоимости товара. В этом случае, кроме проводки Д 90-2 «Себестоимость продаж» К 41-2 «Товары в розничной торговле», следует сделать проводку по отражению суммы реализованной торговой наценки. Это требуется для определения финансового результата от реализации товаров. Проводкой методом «красного сторно» корректируют отраженную по дебету счета 90 «Продажи» покупную стоимость реализованных товаров (в продажной оценке) на разницу между продажной стоимостью реализованных товаров и торговой надбавкой (наценкой), относящейся к проданным товарам. При учете товаров по продажным ценам применяется счет 42 для учета торговой надбавки на поступающие и выбывающие товары. Корректировка дебетового оборота счета 90 и списание со счета 42 торговой надбавки, относящейся к выбывшим (реализованным) товарам, оформляются, как правило, проводкой: Д 90 «Продажи» К 42 «Торговая наценка» — сторно. |

| товароведы как правило оформляют ведомость проданных товаров за период, или за месяц. бухгалтерия списывает сумму согласно их ведомости. |
| …можно придумать какую нибудь среднюю наценку и справкой списать так как написала Милана |
| ваще-то уже тыщу лет, как разрешили не применять 42 счет и вести учет по покупным ценам. 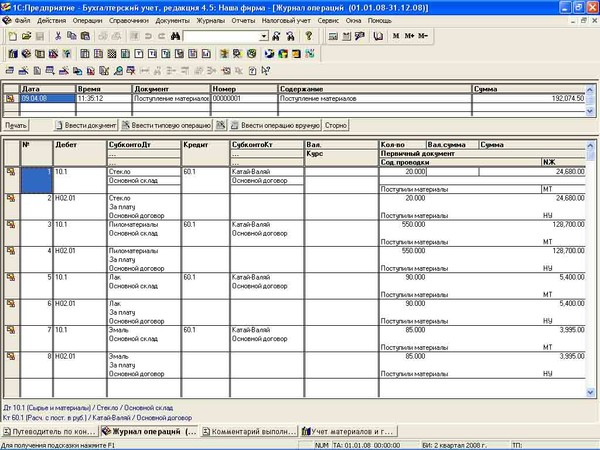 .. у Вас в учетной политике написано, что ведется 42 счет? .. у Вас в учетной политике написано, что ведется 42 счет?откройте Консультант, наберите «розничные продажи 42 счет» и будет Вам счастье.. |
| Дт 41 — Кт 60 — оприходованы приобретенные товары; я бы не рекомендовала ТОРГ-13.. заполнить её проблематично, смысла практически никакого, лучше разработать свою форму межскладской накладной, попроще. |

| ОГРОМНОЕ спасибо всем кто откликнулся!!! |
Рассмотрим, как правильно вести бухгалтерский учет в торговле
Торговля – весьма распространенный и прибыльный вид бизнеса. Рассмотрим, как торговой организации правильно вести бухгалтерский учет в различных ситуациях.
Рассмотрим, как торговой организации правильно вести бухгалтерский учет в различных ситуациях.
Торговля и ее виды
Торговля «вообще» – это перепродажа любых материальных ценностей. Бизнесмен покупает товары, а затем реализует их по более высоким ценам.
Различают следующие виды торговли:
- Оптовая. В этом случае товары обычно продают крупными партиями. Иногда – не очень крупными, но в любом случае это не единичный товар. Как правило, и продавец, и покупатель при оптовых продажах – юридические лица или ИП.
- Розничная. Это продажа товара поштучно или небольшими партиями. В качестве покупателей здесь обычно выступают физические лица.
- Комиссионная. Она осуществляется в рамках, как розничной, так и оптовой торговли. При этом продавец (комиссионер) получает товар у его владельца (комитента) и реализует третьим лицам за вознаграждение.
Бухгалтерский учет в оптовой торговле
Для учета реализуемых ценностей используется счет 41 «Товары».
Поступления товаров от поставщика оформляется в корреспонденции со счетом 60 «Расчеты с поставщиками и подрядчиками».
ДТ 41 – КТ 60
Если бизнесмен работает на общей системе, то на счет 41 попадает сумма без учета НДС, а сам налог отражается на счете 19.
ДТ 19 – КТ 60
Закупка товаров часто связана с дополнительными расходами. Это транспортные услуги, погрузка, для импорта – таможенные платежи и т.п. По общему правилу они должны включаться в себестоимость товара, с помощью той же проводки, что и его закупочная цена.
Однако малые предприятия, которые имеют право вести упрощенный бухгалтерский учет, могут не распределять расходы по доставке на отдельные товары. Они имеют право отнести эти суммы на текущие затраты (п. 13.1 — 13.3 ПБУ 5/01)
Для учета текущих расходов торговые организации используют счет 44 «Расходы на продажу».
Услуги сторонних организаций отражаются проводками:
ДТ 44 – КТ 60 (76)
ДТ 19 – КТ 60 (76) — при работе с НДС
Таким образом нужно учитывать не только расходы по доставке у малых предприятий, но и все иные «внешние» услуги, которые связаны с продажей: реклама, упаковка, страхование и т. п.
п.
Также на счете 44 учитывают и внутренние издержки компании: зарплату, страховые взносы, амортизацию оборудования и т.п.
ДТ 44 – КТ 70 (69,02…)
Реализация в торговле отражается на счете 90 «Продажи» в корреспонденции со счетом 62 «Расчеты с покупателями и заказчиками».
ДТ 62 – КТ 90.1 – товар отгружен
ДТ 90.3 – КТ 68.2 – начислен НДС с реализации
ДТ 90.2 – КТ 41 – списана себестоимость проданных товаров
ДТ 90.2 – КТ 44 – списаны текущие расходы на продажу
ДТ 51 (52,50) – КТ 62 – получена оплата от покупателя
Бухгалтерский учет в розничной торговле
Розничные продавцы имеют право вести учет товаров двумя способами:
- По закупочным ценам
При этом варианте учет поступлений аналогичен оптовой торговле. Каждая единица товара отражается в учете, исходя из цены поставщика и (при необходимости) дополнительных расходов на доставку.
ДТ 41 – КТ 60
ДТ 19 – КТ60
При реализации в розницу вся сумма поступившей за день выручки отражается на 90 счете. Корреспонденция здесь может быть со счетом 50 при продаже за наличные или со счетами 51 или 57 «Переводы в пути» при использовании эквайринга. Необходимость применения счета 57 связана с тем, что многие банки не сразу зачисляют поступившую безналичную выручку на счет бизнесмена.
Корреспонденция здесь может быть со счетом 50 при продаже за наличные или со счетами 51 или 57 «Переводы в пути» при использовании эквайринга. Необходимость применения счета 57 связана с тем, что многие банки не сразу зачисляют поступившую безналичную выручку на счет бизнесмена.
ДТ 50 (51,57) – КТ 90.1 – начислена выручка
ДТ 90.3 – КТ 68.2 – НДС с выручки
ДТ 90.2 – КТ 41 – списана себестоимость проданных товаров.
- По продажным ценам
В отличие от оптовиков, розничные продавцы заранее знают, по какой цене будут реализовывать товар. Поэтому для них возможен учет поступивших товаров по ценам будущей продажи. В этом случае стоимость товара «попадает» на счет 41 уже с учетом наценки. А к двум проводкам по поступлению, рассмотренным выше, добавляется третья, с использованием счета 42 «Торговая наценка».
ДТ 41 – КТ 60
ДТ 19 – КТ60
ДТ 41 – КТ 42
Таким образом, к закупочной цене товаров, которая уже отражена на счете 41, добавляется наценка по ним. Общая сумма наценки по всем приобретенным товарам «собирается» на счете 42.
Общая сумма наценки по всем приобретенным товарам «собирается» на счете 42.
Базовые проводки по реализации и учету себестоимости также будут аналогичны первому варианту. Но в данном случае на 41 счете товары учтены вместе с наценкой, поэтому сумма списанной себестоимости окажется равной сумме реализации.
Чтобы выделить доход бизнесмена, следует уменьшить себестоимость проданных товаров на сумму наценки по ним. Для этого нужна дополнительная проводка по дебету счета 42 и кредиту счета 90.2
Но счет 42 – пассивный, поэтому он не может иметь дебетового сальдо. Поэтому необходимая проводка осуществляется путем «сторнирования», т.е. со знаком «минус». В практике бухгалтерского учета такие операции традиционно выделяют красным.
ДТ 90.2 — КТ 42 (СТОРНО)
Бухгалтерский учет в торговле при порче товара
При любом из видов торговли ценности на складе иногда теряют свой товарный вид. В подобных случаях следует задействовать счет 94 «Недостачи и потери от порчи ценностей».
ДТ 94 – КТ 41 – обнаружена порча товаров
Дальнейшие операции зависят от того, «укладывается» ли сумма потерь в установленные нормы естественной убыли и найден ли виновный.
ДТ 44 – КТ 94 – списание в пределах норм естественной убыли
ДТ 73 – КТ 94 – потери отнесены на виновное лицо
ДТ 91 – КТ 94 – сверхнормативные потери списаны на убытки (если виновный не найден)
Если учет на счете 41 велся с учетом наценки, то списывать ее на убытки по испорченным товарам будет неправильно, так как компания не получила этот доход. Поэтому понадобится еще одна дополнительная проводка.
ДТ 94 – КТ 42 (СТОРНО)
Бухгалтерский учет в комиссионной торговле
Прием товаров на комиссию – это нечто среднее между торговлей и услугами. Комиссионер продает товар комитента и получает от него за это вознаграждение.
- Учет у комиссионера
Комиссионер не становится владельцем продаваемых ценностей, поэтому учитывает их поступление за балансом, по дебету счета 004.
Когда товар реализован, выручка отражается по дебету счета 62 (при оптовой торговле) или счетов 50, 51 (при продаже в розницу). Кредитуется счет 76, в котором нужно выделить отдельный субсчет для расчетов с комитентами.
ДТ 62 (50,51) — КТ 76.1
НДС комиссионер не начисляет. Этот налог платит владелец товаров – комитент, если у него есть такая обязанность.
Одновременно проданные товары нужно списать по кредиту забалансового счета 004.
Договор комиссии может предусматривать, что часть собственных расходов комиссионера на продажу возмещает комитент.
ДТ 76.1 – КТ 60 (10,69,70…)
Остальные затраты, связанные с продажей комиссионных товаров, несет сам комиссионер.
ДТ 44 — КТ 60 (10,69,70…)
Вознаграждение комиссионера отражается в учете, как любое оказание услуг.
ДТ 76.1 – КТ 90.1
Если комиссионер платит НДС, то нужно начислить и этот налог.
ДТ 90.3 – КТ 68.2
Далее нужно списать невозмещенные расходы комиссионера на продажу.
ДТ 90.2 – КТ 44
По итогам сделки комиссионер выплачивает комитенту стоимость проданных товаров, за вычетом своего вознаграждения и возмещаемых расходов.
ДТ 76.1 – КТ 51 (50)
- Учет у комитента
В момент передачи товара комиссионеру его следует отразить на счете 45 «Товары отгруженные».
ДТ 45 – КТ 41
Когда комиссионер продаст товар, комитент должен отразить у себя эту операцию, как обычную реализацию.
ДТ 62 – КТ 90.1
ДТ 90.3 – КТ 68.2
ДТ 90.1 – КТ 45
Вознаграждение комиссионера, с учетом НДС, и его возмещаемые расходы, комитент должен учесть в составе расходов на продажу.
ДТ 44 – КТ 60
ДТ 19 – КТ 60
Вывод
При учете торговых операций важно правильно рассчитать стоимость приобретенных товаров и списать ее в момент реализации.
Варианты отражения этой стоимости зависят от вида торговли и методов учета, применяемых организацией.
Доверьте бухгалтерию команде экспертов
Регулируемые по цвету люминофоры Ca8ZnM (PO4) 7 (M = Lu / Tb, Lu / Eu, Tb / Eu): исследования люминесценции, передачи энергии и термостабильности для белых светодиодов n-UV
Серия Tb 3+ — и Eu 3+ -допированный Ca 8 ZnLu (PO 4 ) 7 (CZLP: Tb и CZLP: Eu 3+ ), а также Ca 8 ZnTb (PO 4 ) 7 : Eu 3+ (CZTP 0003: Eu 3+ ) были получены люминофоры с помощью традиционной высокотемпературной твердотельной реакции.Картины порошковой рентгеновской дифракции (XRD) свежеприготовленных люминофоров показывают, что введение Tb 3+ или Eu 3+ не влияет ни на фазовую примесь, ни на кристаллическую структуру решетки-хозяина CZLP. Спектры фотолюминесценции (ФЛ), зависящие от концентрации, показывают, что даже если Lu 3+ был полностью замещен легирующими добавками, Tb 3+ или Eu 3+ , явления концентрационного тушения не произошло. .Выбросы с регулируемым цветом от зеленого к красному можно реализовать, регулируя тип легирующего иона (Tb 3+ и Eu 3+ ) и их относительную концентрацию. Более того, передача энергии от Tb 3+ к Eu 3+ была подтверждена, и механизм был определен как диполь-квадрупольное взаимодействие. Кроме того, квантовая эффективность составила 0,61, 0,58 и 0,85 для CZTP, CZTP: 0,2Eu 3+ и CaZnEu (PO 4 ) 7 (CZEP) соответственно.В результате было изготовлено устройство на белом светоизлучающем диоде (WLED) с использованием оптимального CZTP: 0,2Eu 3+ , желтый люминофор BaMgAl 10 O 17 : Eu 2+ (BAM: Eu 2+ ) синий люминофор и микросхема ближнего ультрафиолета (n-UV) с длиной волны 370 нм. Полученное устройство отображает подходящий индекс цветопередачи (CRI, ∼81,3) и значение коррелированной цветовой температуры (CCT, ∼2634 K), что указывает на его потенциальное применение в светодиодах n-UV.
У вас есть доступ к этой статье
Подождите, пока мы загрузим ваш контент… Что-то пошло не так. Попробуй еще раз?Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
№ СТОКГОЛЬМ 1.0 # = GF ID NigD_N # = GF AC PF12667.9 # = GF DE NigD-подобный N-концевой OB домен # = GF AU Bateman A; 0000-0002-6982-4660 # = GF GA 24.3 24,3 # = GF NC 24,1 24,2 # = GF TC 24,4 24,4 # = GF PI NigD; # = GF SE JCSG цель Q5LAY5_BACFN # = GF BM hmmbuild HMM.ann SEED.ann # = GF SM hmmsearch -Z 57096847 -E 1000 —cpu 4 HMM pfamseq # = Домен GF TP # = GF CL CL0021 # = GF RN [1] # = GF RM 19531060 # = GF RT Ген, кодирующий нигресцин, продуцируемый Prevotella nigrescens. # = GF RT ATCC 25261. # = GF RA Teanpaisan R, Narawatthana S, Utarabhand P; # = GF RL Lett Appl Microbiol. 2009; 49: 293-298. # = GF DR INTERPRO; IPR024299; # = GF DR SO; 0000417; полипептид_домен; # = GF CC Это семейство белков функционально не охарактеризовано.Этот # = Семейство белков GF CC встречается у видов Bacteroides. Белки в # = GF CC в этом семействе обычно содержится от 234 до 260 аминокислот. # = Длина GF CC. Эти белки содержат N-концевой липопротеин. # = Сайт прикрепления GF CC. Это семейство включает белок NigD, обнаруженный в # = GF CC Nig оперон, кодирующий бактериоцин под названием нигресцин. Она имеет # = GF CC было высказано предположение, что NigD может быть иммунным белком для # = GF CC nigrescin (NigC), потому что он находится непосредственно ниже по течению [1]. Этот # = GF CC домен имеет OB-свертку.# = GF SQ 583 A0A1Y4D0L5.1 / 39-103 IVT.VK … T-TADD..GF … YLQLDDS … TTLLPV..NL.TT … SPFG.-K..K … E.VRALT.NFTPVDQ ….. PT.GGY..TQA ……… VNVNWIDSI F8WXQ8.1 / 38-102 LVT.VK … P-TSGN..SF … FLQLDEK … TTLLPT..NM.TT … SPYG.-N..R … E.VRALV.NYDEVDK ….. PS.QGY..TKA ……… IHINWIDSI B6VVL0.1 / 43-107 LVT.VK … P-VADN..SF … FMQLDDS … TTLLPV..NM.KN … SPYG.EK ..-… E.VRALV.NFDPVNE. …. СС.ГЕЙ..ДКА ……… ВХИНВИДСИ C3RDI3.1 / 43-107 LVT.VK … P-VADN..SF … FMQLDDS … TTLLPV..NM.KN … SPYG.EK ..-… E.VRALV.NFDPVNE….. СС.ГЕЙ..ДКА ……… ВХИНВИДСИ I9QHH7.1 / 43-107 LVT.VK … P-VADN..SF … FMQLDDS … TTLLPV..NM.KN … SPYG.EK ..-… E.VRALV.NFDPVNE. …. СС.ГЕЙ..ДКА ……… ВХИНВИДСИ A0A3A5SRS6.1 / 43-107 LVT.VK … P-VADN..SF … FMQLDDS … TTLLPV..NM.KN … SPYG.EK ..-… E.VRALV.NFDPVNE. …. СС.ГЕЙ..ДКА ……… ВХИНВИДСИ A6L6X9.1 / 43-107 LVT.VK … P-VADN..SF … FMQLDDS … TTLLPV..NM.KN … SPYG.EK ..-… E.VRALV.NFDPVNE. …. СС.ГЕЙ..ДКА ……… ВХИНВИДСИ R9HLY4.1 / 43-107 LVT.VK … P-VADN..SF … FMQLDDS … TTLLPV..NM.KN … SPYG.EK..-… Е.ВРАЛВ.НФДПВНЕ ….. СС.ГЕЙ..ДКА ……… ВХИНВИДСИ A0A069SKQ4.1 / 43-107 LVT.VK … P-VADN..SF … FMQLDDS … TTLLPV..NM.KN … SPYG.EK ..-… E.VRALV.NFDPVNE. …. СС.ГЕЙ..ДКА ……… ВХИНВИДСИ A0A0P0M4L5.1 / 43-107 LVT.VK … P-VADN..SF … FMQLDDS … TTLLPV..NM.KN … SPYG.EK ..-… E.VRALV.NFDPVNE. …. СС.ГЕЙ..ДКА ……… ВХИНВИДСИ A0A1Y4HPQ2.1 / 42-105 LVT.VK … P-TASD..AF … YLQLDDR … TTLLPV..NM.KA … SPFG.-D..K … E.VRALV.NYTNADE ….. PS.GDY..SQA ……… VYVNWIDS- R6SGW7.1 / 42-106 LVT.VK … P-QQDN..SF … YLQLDDS … TTLLPT..NL.AT … SPFG.EK ..-… E.VRALV.NYVETDE ….. AH.GNY..SKA ……… VHINWIDSI R9I029.1 / 43-107 LVT.VK … PLADN — .. SF … FMQLDDS … TTLLPV..NM.KN … SPYG.EK ..-… E.VRALV.NFEQVNE .. … SS.GEY..DKA ……… VHINWIDSI A0A3R6RPM4.1 / 40-104 LVT.VK … P-VSDN..SF … FLQLDDS … TTLLPA..NI.TS … SPFG.-D..K … E.VRALV.NFSEVNE ….. PS.EGY..SKA ……… VHINWIDSI F3PRX2.1 / 43-107 LVT.VK … A-DADN..TL … YLQLDDE … TTLLPT..NL.ST … SPFG.-D..K … E.VRALV.NYTEVEN ….. PS.SKY..SKA ……… VHVNWIDSI S0FDP8.1 / 42-106 LVT.VK … P-QQDN..SF … YLQLDDS … TTLLPT..NL.AT … SPFG.EK ..-… E.VRALV.NYVETDE ….. AH.GNY..SKA ……. ..IHINWIDSI A0A078SW78.1 / 41-105 LVT.VK … TDADQT ..- V … FLQLDDK … TTLLPT..NM.TK … SPYG.EK ..-… E.VRALV.NYKEVDE .. … PS.DIY..TQA ……… VHINWIDSI E5VEY5.1 / 41-105 LVT.VK … TDADQT ..- V … FLQLDDK … TTLLPT..NM.TK … SPYG.EK ..-… E.VRALV.NYKEVDE .. … PS.DIY..TQA ……… VHINWIDSI R7EJS1.1 / 41-105 LVT.VK … TDADQT ..- V … FLQLDDK … TTLLPT..NM.TK … SPYG.EK ..-… E.VRALV.NYKEVDE .. … PS.DIY..TQA ……… VHINWIDSI A0A3A5Z204.1 / 49-113 LVT.ВК … TDADQT ..- V … FLQLDDK … TTLLPT..NM.TK … SPYG.EK ..-… E.VRALV.NYKEVDE ….. PS.DIY..TQA ……… VHINWIDSI A7V7X4.1 / 49-113 LVT.VK … TDADQT ..- V … FLQLDDK … TTLLPT..NM.TK … SPYG.EK ..-… E.VRALV.NYKEVDE .. … PS.DIY..TQA ……… VHINWIDSI R7JBS5.1 / 41-105 LVT.VK … P-TSEN..AF … FLQLDDT … TTLLPV..NM.TS … SPFG.-S..K … E.VRALT.NFSEVNE ….. PS.EGY..SKA ……… VHINWIDSI A0A1M6DV80.1 / 51-115 LVT.VK … SDAEG — .. SM … FLQLDDK … TTLLPT..NV.KK … SPFG.-D..K … E.VRALV.NYDEVDE. …. SS.GDY..NKA ……… VHINWIDSI A0A174V6L2.1 / 41-105 LVT.VK … TDADQT ..- V … FLQLDDK … TTLLPT..NM.TK … SPYG.EK ..-… E.VRALI.NYKEVDE …. .PS.DIY..TQA ……… VHINWIDSI A0A417LLA8.1 / 40-104 LVT.VK … P-VSDN..SF … FLQLDDT … TTLLPA..NI.TS … SPFG.N — .. K … E.VRALV.NFSEVNE ….. PS.EGY..SKA ……… VHINWIDSI A0A3R5ZF05.1 / 40-104 LVT.VK … P-VSDN..SF … FLQLDDS … TTLLPA..NL.TS … SPYG.-D..K … E.VRALV.NFSVVNE ….. PS.EGY..TKA ……… VHVNWIDSI S0GRJ1.1 / 40-104 LVT.VK … P-VSDN..SF … FLQLDDS … TTLLPA..NL.TS … SPYG.-D..K … E.VRALV.NFSVVNE ….. PS.EGY..TKA ………VHVNWIDSI A0A1I4G6B0.1 / 38-102 LVT.VK … A-VDDN..SV … YLQLDDK … TTLLPV..NL.TK … HPFD.N — .. K … E.VRALV.NYKEVDE ….. PS.GTY..SKA ……… VHVNWIDSI A0A4Y8L164.1 / 38-102 LVT.VK … P-INDN..SF … FLQLDDS … TTLLPT..NM.KS … LPFG.-N..K … E.TRALV.NYSETND ….. QK.EGY..SKA ……… VYINWIDSI K9EMJ1.1 / 42-106 LVT.VK … SDADGS ..- L … FLQLDDK … TTLLPT..NV.KK … SPFG.-D..K … E.VRALV.NYDEVDE. …. SS.GDY..SKA ……… IHINWIDSI B3C6h4.1 / 43-107 LVT.VK … G-AADD..AF … FLQLDDK … TTLLPT..NV.KT … SPFG.-D..K … E.VRALV.NYSEVNE ….. PS.GDY..TKA ……… VHINWIDSI R5M2K2.1 / 51-115 LVT.VK … G-AADD..AF … FLQLDDK … TTLLPT..NV.KT … SPFG.-D..K … E.VRALV.NYSEVNE ….. PS.GDY..TKA ……… VHINWIDSI E2NCD8.1 / 43-107 LVT.VK … E-AADG..AF … FLQLDDK … TTLLPT..NV.KT … SPFG.-D..K … E.VRALM.NYSEVDE ….. AS.GEY..TKA ……… VHINWIDSI I8W0L7.1 / 43-107 LVT.VK … E-AADG..AF … FLQLDDK … TTLLPT..NV.KT … SPFG.-D..K … E.VRALM.NYSEVDE ….. AS.GEY..TKA ……… VHINWIDSI R6K869.1 / 43-107 LVT.VK … E-AADG..AF … FLQLDDK … TTLLPT..NV.KT … SPFG.-D..К … Е.ВРАЛМ.НИСЕВДЕ ….. АС.ГЕЙ..ТКА ……… ВХИНВИДСИ R5M504.1 / 40-104 LVT.VK … P-VSGD..GF … YLQLDDS … TTLYPV..NL.KK … SPYG.-D..K … E.VRALV.CFTEVKP ….. VN.DDY..TKS ……… VQINWMDSI E6STG8.1 / 42-106 LVT.VK … TGADRT ..- V … FLQLDDK … TTLLPT..NI.KK … SPYG.EK ..-… E.VRALV.NYKEVNE .. … PS.STC..NKA ……… VYVNWIDSI R7E1F9.1 / 51-115 LVT.VK … G-AADD..AF … FLQLDDK … TTLLPT..NV.KA … SPFG.-D..K … E.VRALV.NYSEVNE ….. PS.GDY..TKA ……… VHINWIDSI A0A415IZA7.1 / 43-107 LVT.VK … G-AADD..AF … FLQLDDK … TTLLPT..NV.KA … SPFG.-D..K … E.VRALV.NYSEVNE ….. PS.GDY..TKA ……… VHINWIDSI A0A0F5IYT2.1 / 39-103 LVT.VK … P-VSDN..SF … FLQLDDT … TTLLPV..NI.AS … SPYG.N — .. K … E.VRALV.NCREVNE ….. PA.EGY..SKA ……… VYVNWIDSI A0A1Y4AJV0.1 / 43-107 IVT.AK … TDTDG — .. TF … FLQLDDS … TTLLPA..NI.AS … SPFG.-D..K … E.VRALL.NFDYAEE. …. SP.GKY..DKA ……… AHVFWIDSI A0A3M8EI80.1 / 41-105 LVT.VK … A-VGDN..SV … YLQLDDK … TTLLPV..NL.TK … HPFD.-S..K … E.VRALV.NYKEVDQ ….. PT.GKY..SKA ……… VHVNWIDSI A0A4S2B2Q1.1 / 49-113 LVT.VK … TDADQT..-V … FLQLDDK … TTLLPT..NM.PK … SPYG.EK ..-… Е.ВРАЛИ.НИКЕВДЕ ….. PG.DIY..TQA …… … ВЬИНВИДСИ A0A1H5WDV9.1 / 41-106 LVT.VK … PNADNS..QF … YMQLDDS … TTLIPT..NL.KS … SPFG.-K..K … E.VRALV.NYYRSNE .. … DA.GHY..SHA ……… VYVNWIDSI A0A373J6L9.1 / 43-107 LVT.VK … N-VDDG..SF … FLQLDDS … TTLLPI..NV.AS … SPFG.EK ..-… E.VRALV.NYDEVDE. …. VS.GRY..SKA ……… VHINWIDSI U6RNR1.1 / 43-107 LVT.VK … N-VDDG..SF … FLQLDDS … TTLLPI..NV.AS … SPFG.EK ..-… E.VRALV.NYDEVDE. …. VS.GRY..SKA ……… VHINWIDSI W4UU32.1 / 44-108 LVT.ВК … S-SIDE..TF … FLQLDDS … TTLLPV..NL.SA … SPYG.-D..K … Е.ВРАЛВ.НЕЕВДК ….. ПС.ГИЙ. .NKA ……… IQINWIDSI Q5LHT5.1 / 43-107 LVT.VK … NATDG — .. SF … FLQLNDS … TTLLPT..NM.SS … SPFG.-D..K … E.VRALV.NYDEVSE. …. SG.GRY..DKA ……… VQVNWIDSI A0A015W2P2.1 / 43-107 LVT.VK … NATDG — .. SF … FLQLNDS … TTLLPT..NM.SS … SPFG.-D..K … E.VRALV.NYDEVSE. …. SG.GRY..DKA ……… VQVNWIDSI R5CN41.1 / 40-104 LVT.VK … P-VSDN..GF … YLQLDDS … TTLFPT..NM.KT … SPYG.-D..K … E.VRALV.CFTEVKP ….. VN.SDY..TKS ……… VEINWMDSI R6N5L8.1 / 43-107 LVT.VK … T-DTDN..TF … YLQLDDN … TTLLPV..NY.SS … SPFG.-D..K … E.VRAIA.NIEEVNE .. … PS.GAY..TKA ……… VQINWIDSI A0A017NLT2.1 / 32-96 LVT.VK … NATDG — .. SF … FLQLNDS … TTLLPT..NM.SS … SPFG.-D..K … E.VRALV.NYDEVSE. …. SG.GRY..DRA ……… VQVNWIDSI A0A015VCA5.1 / 43-107 LVT.VK … NATDG — .. SF … FLQLNDS … TTLLPT..NM.SS … SPFG.-D..K … E.VRALV.NYDEVSE. …. SG.GRY..DKA ……… VQLNWIDSI A0A069S0R5.1 / 43-107 LVT.VK … P-VDDG..SY … FLQLDDS … TTLLPV..NM.TS … SPFG.-Q..K … E.VRALV.NFDETNE ….. СС.ГИЙ..СКА ……… VNINWIDSI A0A496DFV3.1 / 43-107 LVT.VK … P-VDDG..SY … FLQLDDS … TTLLPV..NM.TS … SPFG.-Q..K … E.VRALV.NFDETNE ….. СС.ГИЙ..СКА ……… ВНИНВИДСИ A6LBA3.1 / 43-107 LVT.VK … P-VDDG..SY … FLQLDDS … TTLLPV..NM.TS … SPFG.-Q..K … E.VRALV.NFDETNE ….. СС.ГИЙ..СКА ……… ВНИНВИДСИ R6FEG5.1 / 43-107 LVT.VK … H-ASDN..LL … FLQLDDS … TTLLPC..NV.KT … SPFG.EK ..-… E.VRALV.NYTPVDE. …. PS.GEF..DQA ……… IHINWIDSI A0A4Y8L5N2.1 / 39-103 LVT.VK … Q-SSDK..TV … FLQLDDK … TTLLPV..NV.KS … SPFG.N — .. K … E.VRALV.NYKEVNE ….. СН.КЕЙ..ТКА ……… ВЬИНВИДСИ A0A3R6HN16.1 / 42-106 -AT.VK … P-TSST..SY … YIQLDDS … TTLLPV..DG.SI … PYFG.LD..E … E.RRALV.NFTLLSD ….. СИ.КГЫ..ША ……… ВКВНКИДСИ B3JI21.1 / 51-115 LVT.VK … S-DAND..SF … FLQLDDS … TTLLPV..NL.AS … SPFG.-D..K … E.VRALA.NIEEVDE ….. PS.GAY..TKA ……… VHINWIDSI R6CE01.1 / 51-115 LVT.VK … S-DAND..SF … FLQLDDS … TTLLPV..NL.AS … SPFG.-D..K … E.VRALA.NIEEVDE ….. PS.GAY..TKA ……… VHINWIDSI R7A5N4.1 / 43-107 LVT.VK … L-SADN..TP … YLQLDDS … TTLRPV..NM.TT … SPFG.-D..K … E.VRALT.NFDFSEE ….. AP.QGY..DKA ……… VFVNWIDSI A0A4Q0TW68.1 / 43-107 LVT.VK … NATDG — .. SC … FLQLNDS … TTLLPT..NM.TS … SPFG.EK ..-… E.VRALV.NYDELDE .. … SG.GKY..DKA ……… IQVNWIDSI E4VXD8.1 / 43-107 LVT.VK … NATDG — .. SC … FLQLNDS … TTLLPT..NM.TS … SPFG.EK ..-… E.VRALV.NYDELDE .. … SG.GKY..DKA ……… IQVNWIDSI R5R722.1 / 43-107 LVT.VK … NATDG — .. SC … FLQLNDS … TTLLPT..NM.TS … SPFG.EK ..-… E.VRALV.NYDELDE .. … SG.GKY..DKA ……… IQVNWIDSI A0A415DSB3.1 / 42-106 LVT.VK … P-IDNK..SF … FLQLDDS … TTLLPV..NV.TS … SPYG.-D..R … E.VRALV.NCQLVNE ….. PS.EGY..DKA ……… VHINWIDSI G8UNM9.1 / 39-105 LAT.VN … PLDNGR..IY … SLTLDDG … KTLWPA..AT.RA … PYYK.PK..A … N.QRAFV.YYTILSD … ..SF.QGY..NHA ……… VQIHDIRNI E6K5D1.1 / 38-103 LVT.LK … TNPTDG..TF … FMQLDDS … TTILPT..NM.KT … SPYG.N — .. K … E.VRALA.NFKIGEG .. … QS.GHY..SKS ……… AYVNWIDSI R6DE06.1 / 41-105 LVT.VK … T-EADK..TV … YLQLDDK … TTLFPT..NM.KQ … SPFG.-D..K … E.VRALV.NCVATGG ….. NS.AKY..DKA ……… VYVNWIDSI F9YT36.1 / 38-101 -VT.VR … PQADG-..SF … FLQLDDV … TKLTPT..NM.KK … SPFG.N — .. K … E.VRALV.NYTKENS ….. NT.KSK..SQD …… … VRINWIDSI A0A4Y8V811.1 / 38-102 LVT.VY … P-TAPE..GF … FMQLDES … TSLVPT..NM.KT … SPFG.-D..K … K.VRALV.NYTIEER ….. SY.GGN..QQS ……… VYVNWIDSI A0A1G4GAL4.1 / 37-101 LVT.VK … H-AGDK..TV … FLQLDDK … TTLLPV..NL.TT … SPFG.EK ..-… E.VRALV.NYKEVDI. …. PS.SGY..DQA ……… VHVSWIDSI A0A3N2MGT2.1 / 39-102 LVT.VC … P-TSDG..GF … IMQLDNN … TTLYPT..NM.DK … SPYG.-S..K … E.VRALV.NYDEDNL …..- E.KSD..RKD ……… VHVNWIDSI A0A1Y3X7L0.1 / 41-105 IVT.AK … TDTDG — .. TF … YLQLDDS … TTVLPV..NI.AT … SPFG.-D..K … E.VRALM.NFDFSEE ….. SA.GKY .. DQA ……… VTVYWMDSV A0A1Y4DWD7.1 / 41-105 IVT.AK … TDTDG — .. TF … YLQLDDS … TTVLPV..NI.AT … SPFG.-D..K … E.VRALM.NFDFSEE. …. SA.GKY..DQA ……… VTVYWMDSV A0A1Y4ELM0.1 / 41-105 IVT.AK … TDTDG — .. TF … YLQLDDS … TTVLPV..NI.AT … SPFG.-D..K … E.VRALM.NFDFSEE. …. SA.GKY..DQA ……… VTVYWMDSV A0A1R3T251.1 / 37-101 LVT.VK … L-IDDN..SV … YLQLDDN … TTLLPV..NL.TK … HPFD.-S..K … E.VRALV.NYKEVDE ….. PS.GIY..SKA ……… VHVNWIDSI R5TT48.1 / 58-122 IVT.AK … TDTDG — .. TF … YLQLDDS … TTVLPV..NI.AT … SPFG.-D..K … E.VRALM.NFDFSEE … ..SA.GKY..DQA ……… VTVYWMDSV S8GQZ3.1 / 7-73 LVT.VK … PSIDNS..SF … YLQLDDN … TTLAPT..NM.KV … SPFG.-K..K … E.VRALV.NYTLDDQ .. … AE.TSP.hSRA ……… VRINWIDSI E7RPX0.1 / 38-103 LVT.VK … PNSDNS..SF … YMQLNDS … TTLLPA..NM.KA … SPFG.-K..K … E.VRALV.NYTMVDR .. … PT.MHS..GRA ……… VNINWIDSI A0A3E4N7W9.1 / 44-108 LVT.VK … Q-SADN..TP … YLQLDDS … TTLRPV..NM.AS … SPFG.EK ..-… E.VRALA.NIELADE. …. AP.QEY..DKA ……… VFINWIDSI R5KUX9.1 / 40-104 LVT.VK … P-IEGA..GF … YMQLDDS … TTLYPV..NM.AA … SPYG.-D..K … E.VRALV.CYREVNP ….. VN.DYY..SKS ……… VEVSWMDSI R6X446.1 / 38-103 LVT.LK … TNAETG..AF … YMQLDDA … TTLVPT..NI.KT … APYG.GK ..-… E.VRALV.NFKEDQK … ..ST.GHN..SRS ……… VYVNWIDTI R5W3M0.1 / 44-108 LVT.VK … Q-SADN..TP … YLQLDDS … TTLRPV..NM.TT … SPFG.EK ..-… E.VRALA.NIELADE. …. AP.QEY..DKA ……… VFINWIDSI R6EEJ0.1 / 40-102 MVT.VK … TANDGD ..- W … YMQLDDE … TTLYAT..NM.HQ … APYD .— .. K … E.VRALV.NCYEVRG….. DA.NGY..TKA ……… VYVNWIDS- U2L0C3.1 / 38-103 IVT.VK … PATAKT..PF … YLQLDDS … TVLRPV..NM.TK … SPYG.-D..K … E.VRAFV.NFSVTEA .. … GH.NKR..DYP ……… VRVNWIDSI R5P4T4.1 / 38-101 LVT.VY … P-QGAN..GF … YMQLDSV … TTLVPT..NL.KS … SPFG.EK ..-… T.VRALV.NYNEESG. ….- S.NGG..VKN ……… VKVNWIDSI F9ZB83.1 / 38-102 LVT.VY … P-SAPT..GF … FMQLDES … MSLVPT..NM.KA … SPFG.-D..K … K.VRALV.NYTIEEE ….. SY.GGN..QLS ……… VYVNWIDSI R5MHV3.1 / 43-107 LVT.AK … T-DTDN..TF … FLQLDDS … TTLLPV..NY.TT … SPFG.EK..-… Е.ВРАЯ.НИЕЕВНЕ ….. АС.ГАЙ..ТКА ……… ВРИНВИДСИ L1N4R3.1 / 38-103 LVT.VK … PNVNNT..SF … SMQLDDS … TVLHPV..NR.KT … SPFG.TK ..-… E.VRALV.NYTVVDR … ..PT.LGK..GIN ……… VNVNWIDSI A0A374TFI8.1 / 41-104 LVT.VK … Q — NAD..AC … YLQLDDN … TTLLAK..NL.KT … PVFD.GK ..-… E.VRALV.NYTATDE ….. KS.DKY..SQV ……… VHVNWIDSI B5CTS2.1 / 44-108 LVT.VK … Q-SADN..TP … YLQLDDS … TTLRPV..NM.TT … SPFG.EK ..-… E.VRALA.NIELADA. …. AP.QEY..DKA ……… VFINWIDSI A0A415VK38.1 / 41-104 LVT.VK … Q — NAD..AC … YLQLDDN … TTLLAK..NL.KT … SVFD.GK ..-… E.VRALV.NYTATDE ….. KS.DKY..SQV ……… VHVNWIDSI B0NMY5.1 / 38-104 IGT.VR … I-IDGK..DY … YFALDEG … TKMFPG..DT.AQ..vDNYT.LV..E … G.QRAFV.YFNLLDE .. … EV.TGY..DYN ……… AKINHVENI A0A108TD91.1 / 38-104 IGT.VR … I-IDGK..DY … YFALDEG … TKMFPG..DT.AQ..vDNYT.LV..E … G.QRAFV.YFNLLDE .. … EV.TGY..DYN ……… AKINHVENI S3YC02.1 / 38-104 IGT.VR … I-IDGK..DY … YFALDEG … TKMFPG..DT.AQ..vDNYT.LV..E … G.QRAFV.YFNLLDE .. … EV.TGY..DYN ……… AKINHVENI A0A098BYA6.1 / 38-102 LVT.VK … E-TADD..KL … FFQLDDQ … TTLLPV..NI.NT … HPYG.GK ..-… E.VRALV.NYKEVDV ….. PS.GDF..DKA ……. ..VHVNWIDSI R5CTH7.1 / 39-106 LVT.VK … P-MDEG..SF … CMQLDDS … TVVYPY..NL.KK … SPFG.-D..K … E.VRALA.NLTPVDAg ..teSD.ANY..MRY ……… VVVNWIDSI A0A1C2BRD2.1 / 39-104 IAT.VE … P-LDNN..TY … SLILDDG … TKLWPA..AT.NY … PGYD.PK..N … D.QRALV.NYTILSD. …. SI.QGY..DHF ……… IKVNSIQNI A0A3L8A5Q7.1 / 42-104 —.- M … S-QDSK..DF … YFTLDDG … KKMYPS..NG.EG..wSNAD.FA..D … G.QRAFV. MFNELEE ….. PV.GGY..DYN ……… VQVKQIKEI A0A229I8I4.1 / 41-104 LVT.ВК … Q — НАД..АС … YLQLDDN … TTLLAK..NL.KT … PVFD.GK ..-… E.VRALV.NYTATDE ….. KS.DKY. .SQV ……… VNVNWIDSI A0A6H0KPM2.1 / 42-104 —.- M … S-QDSK..DF … YFTLDDG … KKMYPS..NG.QG..wSNAD.FV..D … G.QRAFV. MFNELEE ….. PV.GGY..DYN ……… VQVKQIKEI D1PBP8.1 / 41-104 LVT.VK … K — SGD..AC … YLQLDDN … TTLLAK..NL.K -… PTIF.DG..K … E.VRALV. NYTQTSE ….. KS.EKY..NQV ……… IHVNWIDSI R6BSA4.1 / 41-104 LVT.VK … K — SGD..AC … YLQLDDN … TTLLAK..NL.K -… PTIF.DG..K … E.VRALV. NYTQTSE ….. KS.EKY..NQV ……… IHVNWIDSI A0A3R6B2E2.1 / 41-104 LVT.VK … Q — NAD..AC … YLQLDDN … TTLLAK..NL.KT … SVFD.GK ..-… E.VRALV.NYTATDE .. … KS.DKY..SQV ……… VYVNWIDSI A0A5C6HLR4.1 / 40-100 —T.VR … V — TGN..AF … YLVSDTW … GTLWPV..NI.DL … GWYQ.PV..D … G.QRVLT .VFNPLYD ….. NY.QGY..DHA ……… VKLLSIR— R6YHQ9.1 / 37-102 WVT.VH … V-EGTG..AY … SFTGDTW … GTMWPA..AT.AI … WGYR.PV..E … G.QRAIL.YFNPLYE. …. NF.QSY..DYG ……… IKPEYIYDI A0A374UUF0.1 / 38-104 IGT.VR … I-VDGK..DY … YFALDEG … TKMFPG..DT.AQ..iNNYT.LI..E … G.QRAFV.YFNLLDE .. … КВ.НГИ..ДЫН ………AKINHVENI R6A567.1 / 41-106 LVT.VQ … PEGNGK..PL … TLKLDDK … TTLTPT..NL.TA … SPFG.N — .. K … V.VRAIC.NLTFEEN .. … PT.DKY..NQK ……… ARVNWIDSI A0A017NHH0.1 / 37-98 WAT.VR … V-VGGD..TY … SLNADRW … GTLWPA..AT.AI … PFYK.PI..D … G.QRVIT.YFNPLYD. …. NY.EGY..DHA ……… VKVEH —- Q5LHD5.1 / 37-98 WAT.VR … V-VGGD..TY … SLNADRW … GTLWPA..AT.AI … PFYK.PI..D … G.QRVIT.YFNPLYD. …. NY.EGY..DHA ……… VKVEH —- A0A015U643.1 / 37-98 WAT.VR … V-VGGD..TY … SLNADRW … GTLWPA..AT.AI … PFYK.PI..D … G.QRVIT.YFNPLYD….. NY.EGY..DHA ……… VKVEH —- A0A015VI61.1 / 37-98 WAT.VR … V-VGGD..TY … SLNADRW … GTLWPA..AT.AI … PFYK.PI..D … G.QRVIT.YFNPLYD. …. NY.EGY..DHA ……… VKVEH —- R6XAW1.1 / 40-103 LVT.VK … P — ISD..GC … YFQLDDN … TTLKPG..NI.NK … KPFG.-D..K … E.VRALV. NYSVTDQ ….. KD.ETY..GQV ……… VNVNWIDSI R6LGW7.1 / 38-104 IGT.VR … I-IDGK..DY … YFALDEG … TRMFPG..DT.AQ..vNNYT.LV..E … G.QRAFV.YFNLLDE .. … К.В.ДГЫ..ДЫН ……… АКИНХВЕНИ A0A1Y4JT44.1 / 38-104 IGT.VR … I-IDGK..DY … YFALDEG … TRMFPG..DT.AQ..vNNYT.LV..E … G.QRAFV.YFNLLDE ….. KV.DGY..DYN ……… AKINHVENI R6DCV4.1 / 44-104 —. — …— QDSK..DF … YFTLDDG … KKMYPS..NG.QG..wNGAD.FA..D … G.QRAFV. MFNELEE ….. PV.GGY..DYN ……… VQVKQIKEI R6G7I7.1 / 36-102 IGT.IK … V-VDGQ..EY … YFALDEG … TKMYPG..DT.TN..iHNYT.VV..P … E.QRAFV.YFDLLEE .. … KV.SGY..DYN ……… AAIYRIENI A0A069S6h4.1 / 40-108 -AT.VV … P-EGNN..VY … YLRLDDG … DKLWPA..AT.NY … PNYQ.PK..P … N.QRALV.NFTILSDst .qsNL.GGF..SHY ……… ВКВНАИХНИ R5PC11.1 / 41-106 IVT.VK … PETAES..KF … FLQLDDS … TTLTPL..NM.NS … SPYG.-S..K … E.VRAFI.NFRYAEQ ….. QN.NKR..NFN ……… VYVNWMDSI A0A069CZF4.1 / 39-103 —T.IK … V-KAGQ..DY … YFALDDG … KKMLPG..DT.TS..iNNYL.VK..D … G.QRAFV.YFNLLEE ….. KK.EGY..DYN ……… AKVQYIENI A0A380YJR9.1 / 39-105 IGT.VR … I-VDGK..DY … YFALDEG … TKMFPG..DT.TR..vNNYA.LV..E … G.QRAFV.YFNLLDE .. … KV.DGY..DYN ……… AQINHVENI E5X2B0.1 / 39-105 IGT.VR … I-VDGK..DY … YFALDEG … TKMFPG..DT.TR..vNNYA.LV..E … G.QRAFV.YFNLLDE .. … KV.DGY..DYN ……… AQINHVENI A0A0B7H0Z0.1 / 38-101 -VT.VR … P-QTDD..TF … FLQLDDV … TRLNPT..NM.KK … SPFG.-K..K … E.VRALV.KYTKESN ….. DT.KSK..AQN …… … ВРИХВИДСИ R6EWA2.1 / 38-101 LVT.VY … P-VTSE..SF … RMQLDDQ … TVLFPT..NM.KA … SPFG.EK ..-… K.VRALV.SYEYEEG. ….- A.TGA..QRN ……… VRVNWIDSI R5FE74.1 / 40-102 IVT.VK … TAENGD ..- W … YMQLDDS … TTLYAT..NI.HQ … APYN .— .. K … E.VRALVsCYETGGD … ..- A.NGY..SKA ……… VFVNQIDS- A0A2V3PJZ6.1 / 38-104 IAT.VN..sIDSTAG..TY … YLTLDNG … ETLWPV..SS.A -… PHFV.PK..S … N.QRVLV.NFTLLSD … ..KI.GEY..DHY ……… ARINQIQNI A0A250FWM2.1 / 39-102 -VT.VR … PQSDG — .. SF … FLQLDDV … TRLNPT..NM.KK … SPFK.-D..K … E.VRALV.NYTKEKT ….. DT.KQK .. IEN ……… VHINWIDSI A0A496DMC6.1 / 40-108 -AT.VV … P-EGNN..VY … YLRLDDG … DKLWPA..AT.NY … PNYQ.PK..P … N.QRALV.NFTILADst .qsNL.GGF..SHY ……… ВКВНАИХНИ R7DVR5.1 / 40-100 -AT.IR … V — KGN..TF … YMESDTW … GTLWPI..NT.DM … GWYQ.PV..D … G.QRVVT. VFNPIYD ….. NY.EGY..DHA ……… VKILRL — R5M8N4.1 / 40-100 -AT.IR … V — KGN..AF … YMESDTW … GTLWPI..NT.DM … GWYQ.PV..D … G.QRVVT. ВФНПИЙД ….. НЙ.ЭГИ..ДХА ……… ВКИЛРЛ — A0A415IG08.1 / 40-100 -AT.IR … V — KGN..TF … YMESDTW … GTLWPI..NT.DM … GWYQ.PV..D … G.QRVVT.VFNPIYD. …. NY.EGY..DHA ……… VKILRL — B3CGK2.1 / 40-100 -AT.IR … V — KGN..AF … YMESDTW … GTLWPI..NT.DM … GWYQ.PV..D … G.QRVVT. VFNPIYD ….. NY.EGY..DHA ……… VKILRL — E2NFC3.1 / 41-100 —T.VR … V — KGN..TF … YLESDTW … GTLWPI..NT.NM … GWYQ.PV..D … G.QRIIT .IFNPIYD ….. NY.EGY..DHA ……… VKIEKI — I8WNN0.1 / 41-100 —T.VR … V — KGN..TF … YLESDTW … GTLWPI..NT.NM … GWYQ.PV..D … G.QRIIT .IFNPIYD ….. NY.EGY..DHA ………VKIEKI — R6KJ61.1 / 41-100 —T.VR … V — KGN..TF … YLESDTW … GTLWPI..NT.NM … GWYQ.PV..D … G.QRIIT .IFNPIYD ….. NY.EGY..DHA ……… VKIEKI — R5C043.1 / 38-102 WAT.VR … V — GGG..AF … YLDCDVW … GTLWPV..NT.DM … GWYT.PV..D … G.QRVIT.YFNPLWD ….. NY.EGY..DHA ……… VKILGIQNV A0A401LX21.1 / 37-102 —T.IN … TIQDSK..DF … YFTLDDG … KTMFPS..NP.QG..mGAKE.YE..D … G.QRAFV.IFNKLDD .. … PV.NGY..DYN ……… IQVKRIQEI A0A2N4S6A4.1 / 38-101 IAT.VE … TDDNSS..AY … WFILDDG … TTLWPA..AS.DI …- QYL.PK..Q … G.QRIIL.NYSILSN….. QK.NEF..DHY ……… IKINYIW— A6LF92.1 / 40-108 -AT.VV … P-EGNN..VY … YLRLDDG … DKLWPA..AT.NY … PNYQ.PK..P … N.QRALV.NFTILADst .qsNL.GGF..SHY ……… ИКВНАИХНИ R7DLR7.1 / 42-105 —D.MV … V-KEGE..SP … YLQLDSK … KTMLPS..NT.SI … PSFF.LE..D … G.RRVII. NFSLLQE ….. NY.MGY..DYY ……… VAINDIDTV A0A5C6HL69.1 / 40-106 IGT.LK … IIA-GK..DY … YFALDNG … KKMYPG..DT.TY..iHNYT.PV..D … N.QRVFI.YFQPLKQ .. … PA.TGY..DYN ……… VQLYRLENI A0A174JRQ3.1 / 43-104 —. — …- SEDSK..EF … YFTLDNG … KTMFPS..NS.QA..wGGEK.FE..N … G.QRAFV.IFNELEQ ….. PV.NGY..DYN ……… IQVRDITKV U2P385.1 / 32-96 LVT.LK … T-NASG..TF … FLQLDDS … TTVIPT..NM.KA … SPYG.-K..K … E.VRALA.NINMDAK ….. EP.KDH..VKS ……… GYVNWLDSI K9ER73.1 / 40-105 -GT.LE … I-IEGR..EY … FFNLDDG … DKMYPS..DT.TY..iHNYE.LV..D … G.QRVFV.QFLPLEE. …. DK.PGY..EYN ……… VKLIQLQNI A0A374CCL3.1 / 40-103 LVT.VK … V — TGD..RC … YFQLDDQ … TTLAPG..NI.SK … KPFG.-D..K … E.VRALV. NYTMTDR ….. KD.DKY..GAV ……… VNVNWIDSI R6BGM4.1 / 40-103 LVT.VK … V — TGD..RC … YFQLDDQ … TTLAPG..NI.SK … KPFG.-D..K … E.VRALV.NYTMTDR ….. KD.DKY..GAV ……… VNVNWIDSI A0A4Y8VNM2.1 / 40-103 LVT.VK … V — TGD..RC … YFQLDDQ … TTLAPG..NI.SK … KPFG.-D..K … E.VRALV. NYTMTDR ….. KD.DKY..GAV ……… VNVNWIDSI R5P1C2.1 / 42-105 LVT.VK … GGTDS -..- C … FLQLDDE … TTLWPV..NL.SS … SPFG.EK ..-… E.VRALV.YFTDVDG. …. EK.HFC..TRA ……… VYVNWMDSI A0A1Y4GR83.1 / 41-104 —. MV … V-EEGH..SP … YIQLDDK … KTMFPS..NG.DR..lPSYL.TK..N … G.KRVIV.NFTFLDG ….. QR.EGY..DYY ……… IVVNDIDSV Q89YL2.1 / 44-104 —. — …— ПДСК..DF … YFTLDDG … KTMFPS..NG.NG..wISDK.NK..D … G.QRAFV.IFKELEE ….. PV.KGY..DYN …….. .IQVREIKEI R7J4Y2.1 / 71-133 LVT.VV … P-QDGG..SF … VMNLDDA … TVLIPT..NM.SR … SPYG.-D..K … E.VRALV.NYIDEDS …..—. RAA..ERR ……… VKVVWIDSI A0A4Z0V888.1 / 37-102 IVT.VC … P-EEDG..SF … VMNLDNE … TVLQPT..NM.NK … SPFG.-K..K … E.VRALV.NYTETVA ….. SE.GKS.vLRN ……… VQINWIDSI K9DZ78.1 / 39-100 WAT.VR … V — TGN..AF … YLNSDTW … GTLWPI..NT.DM … GWYQ.PV..D … G.QRVVT.VFNPIYD ….. NY.GEY..DHA ……… VKIERI — A0A374VA37.1 / 39-104-GT.IR … I-IEGK..DY … YFSLDDG … SKMYPS..DT.AY..iNNYA.LT..E … G.QRAFI.YFNLLDE ….. KM.DGY..DYN ……… АКИНХИЕНИ A0A1Y3WD59.1 / 39-104 -GD.MV … V-EEGS..DP … YIQMDDK … KTLFPS..NG.DR..lPSYL.KK..N … G.QRVIV.NFTFLDE. …. HR.EGY..DYY ……… ILVNDIDSV A0A4S2AES5.1 / 44-108 WAT.VR … V — TGN..AF … YLNCDVW … GTLWPV..NT.DI … GWYD.AV..D … G.QRVIT.VFNPLQD ….. NY.AGY..DHA ……… VKLLRLQNI S3YBA8.1 / 38-102 WAT.VR … V — TGN..AF … YLDCDVW … GTLWPV..NT.DL … WWYE.PV..D … G.KRVIT.MFNPLSD ….. EF.DGY..DHA ……… VKILSLQDV A0A120A394.1 / 38-102 WAT.VR … V — TGN..AF … YLDCDVW … GTLWPV..NT.DL … WWYE.PV..D … G.KRVIT.MFNPLSD .. … EF.DGY..DHA ……… VKILSLQDV B0NV25.1 / 38-102 WAT.VR … V — TGN..AF … YLDCDVW … GTLWPV..NT.DL … WWYE.PV..D … G.KRVIT.MFNPLSD ….. EF.DGY..DHA ……… VKILSLQDV A0A1h3W4X3.1 / 43-103 —. — …— PDSK..DF … YFTLDDG … KTMFPS..NG.NGw.iSD-K.SK..D … G.QRAFV .IFKELEE ….. PV.KGY..DYN ……… IQVREIKEI A0A1Y3R2P4.1 / 22-88 FAT.VK … PLENND ..- F … FVVLDDG … KTVYPG..DK.SR..iGSYK.AE..E … G.QRVII.YFNLLNN … ..KV.EGY..DYN ………MALYYIQDI W0EM50.1 / 40-104 —D.MV … V-AEDH..DP … YILLDNK … KTMFPS..NG.TR..lPNYL.TK..D … G.QRVII.NFSYLDG ….. NR.EGY..DYY ……… ILVNDIDSV A0A1M5CEY4.1 / 37-103 LAT.VN..pIDTETG..TY … YLTLDNG … ATLWPA..AS.NV …- NYS.PK..A … N.QRVVV.NYTLLSD … ..QV.GEY..DHY ……… VKINGLDEI R5S0V2.1 / 40-107 -GT.IQ … TLSQDE.gDF … YVLLDDN … SKLFPG..DM.SS..lRGYE.AT..D … G.QRAFV.FFDELPE …. .EM.PGY..DYN ……… AKIRWIQDI E0NSI9.1 / 41-106 LVT.VK … PNIDNS..AF … YMQLNDE … ETLLAT..NI.KT … SPFG.-K..K … E.VRALV.NYTLERR….. KD.PHY..SHL ……… VWVNWIDSI R5EVT2.1 / 42-104 —.- V … V-KDGS..EP … YIQLDSK … QTLFPS..NG.SR..lPNYL.LK..D … G.KRVIV. NFSFLEG ….. QR.EGY..DYY ……… ILINDIDSV F3QTF3.1 / 42-105 LVT.VK … GGTDS -..- C … FLQLDDE … TTLWPV..NL.SS … SPFG.EK ..-… E.VRALV.HFTDVDE. …. DR.HSR..ARA ……… VYVNWMDSI A0A4Y1WYW5.1 / 35-103 -VTiLN … PGGLSG..TF … YFKADNG … QTLYPS..QF.AVp.tGGYN.PE..D … G.QRAFI.YYSLLEE ….. KS .QQY..DYN ……… IKLYGISEI R7PCB7.1 / 38-102 LVT.VK … H-TADN..VC … YLQLDDS … TTLFPT..NL.SR … SPFG.-Q..K … E.VRALA.DLVMTGN ….. GR.DGF..TQQ ……… AYINWIDSI A0A098C1J3.1 / 37-100 -AT.II … N-EGDN..AY … SLQLDNG … KRLWPA..AK.AV … NFLP.TN ..-… H.QRVLV.NYTILSD ….. QK.DGY..DHY ……… VKINDIWKI A0A173WIU3.1 / 46-106 —. — …— QDSK..DF … YFTLDNG … EKMYPN..NG.QE..wNSAE.FA..E … G.QRAFV. IFNELET ….. PV.NGY..DYN ……… VQVKQINEI C9KY10.1 / 46-106 —. — …— QDSK..DF … YFTLDNG … EKMYPN..NG.QE..wNSAE.FA..E … G.QRAFV. IFNELET ….. PV.NGY..DYN ……… VQVKQINEI K5BU75.1 / 46-106 —. — …— QDSK..DF … YFTLDNG … EKMYPN..NG.QE..wNSAE.FA..E … G.QRAFV.IFNELET ….. PV.NGY..DYN ……… VQVKQINEI A0A3N2N7Z4.1 / 39-103 LVT.VK … T-SPDK..TP … YLQLDEK … TTLLPT..NL.KT … GLFD.G — .. K … Q.VRALA.NISITDQ ….. PA.GDY..SKA ……… AYVNWIDSI U2BVA1.1 / 42-104 —.- S … D-AEKN..RF … YLTQDNG … QTLFPS..DI.RY..lGNYK.PV..D … G.QRAFV. TCNFLDE ….. KA.AGY..DYN ……… IRILKIVNI W4PTV4.1 / 42-104 —.- S … D-AEKN..RF … YLTQDNG … QTLFPS..DI.RY..lGNYK.PV..D … G.QRAFV. TCNFLDE ….. KA.AGY..DYN ……… IRILKIVNI A0A4P7W3N5.1 / 43-104 -VT.VC … PAADG-..KF … SMVLDDE … TSLVAT..NM.TS … SPFG.-D..K … E.VRALV.NYTEDSS …..- Y.NGG ..- KG …. ….. ВЬИНВИДСИ A0A3N2KVY8.1 / 43-104 -VT.VC … PAADG — .. KF … SMVLDDE … TSLVAT..NM.TS … SPFG.-D..K … E.VRALV.NYTEDSS …..- Y.NGG ..- KG ……… VYINWIDSI R5S8X9.1 / 38-102 WAT.VR … V — GGG..AF … YLDCDVW … GTLWPV..NT.DL … GWYK.PV..D … G.QRVIT.YFNPLWD ….. DF.EGY..DHA ……… VKILGIQNV A0A1M5A4H5.1 / 35-102 IGT.IN … TIQDTK..DL … YFTLDDG … KTMFPS..NP.QE..mGAKE.YE..D … G.QRAFI.IFNKLDE …. .PV.NGY..NYN ……… IQVKRIQEI Р6ММ70.1 / 37-101 WVT.VR … V-EGEG..VY … SFTGDTW … GTMYPA..VN.GV … WNYS.LV..E … G.QRAFL.VFNPLFD ….. NY.YGY .. DVG ……… IKAEQIYP- A0A562VF25.1 / 38-100 WAT.VR … V — TGN..AF … YLNCDTW … GTLWPV..NT.DL … GWYE.AA..D … G.QRVLT.VFNPLWD ….. NY.QGY..DHA ……… VKLLKLR— R5C5K4.1 / 38-105 -GT.IK … TLSDSE..GD..fYVLLDED … SKLFPG..DM.SN..lHNYK.AV..D … G.QRAFV.FFDELPE …. .EM.PGY..EYN ……… AKIRWIEDI A0A419W8X1.1 / 37-102 -GT.VV … K — SSD..YF … SVTTDGG … DKLWPS..AT.NV … DPEH.LE..D … G.MRVFV. НИТИЛДДА … eES.SGY..DYY ……… VKINSMSEI F3PQN3.1 / 39-105 IGT.IK … VIEEND ..- Y … YFTLDEG … SKLYPG..DT.TA..lYNYT.VV..D … G.QRAFV.YFDELSE ….. КК.ЕГИ..ЭЙН ……… АКИХИЕНИ A0A4Q0U563.1 / 37-97 WAT.VR … V-VGGD..TY … ALNADHW … GSLWPA..AT.AI … PGYR.PI..D … G.QRVIT.YFNPLYD. …. NY.EGY..DHG ……… VKVE —— E4VTV5.1 / 37-97 WAT.VR … V-VGGD..TY … ALNADHW … GSLWPA..AT.AI … PGYR.PI..D … G.QRVIT.YFNPLYD. …. NY.EGY..DHG ……… VKVE —— R5RI19.1 / 37-97 WAT.VR … V-VGGD..TY … ALNADHW … GSLWPA..AT.AI … PGYR.PI..D … G.QRVIT.YFNPLYD. …. NY.EGY..DHG ……… ВКВЕ —— A0A553ENF9.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NA.QG..wNNKD.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DHN ……… IQVRGINPI A0A3P2ABN0.1 / 38-102 WAT.VR … V — TGN..AF … YLNCDTW … GTLWPV..NV.DL … GWYD.AV..D … G.QRVLT.VFNPLWD ….. NY.EGY..DHA ……… VKLLKLQNV A0A6F9Z8W2.1 / 37-103 WAT.VN … V-KGAH..VY … DFTGDRW … GKIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD. …. NYpEGY..DCS ……… VKVLNIKEI R9IAV6.1 / 37-103 WAT.VN … V-KGAH..VY … DFTGDRW … GKIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD….. NYpEGY..DCS ……… VKVLNIKEI R6E574.1 / 38-95 -VT.VK … P — DGD..KF … YLQLDDK … TVIYPS..NM.TK … SPYG.-D..K … E.VRAFT .NYKLESE …..- S.NG — .. VQI ……… GNVLWL — A0A4S4HBE6.1 / 38-103 LVT.VS … PGSDG — .. TF … VMYLDDN … TVLTPS..NM.KS … SPFK.EK ..-… E.VRALV.NYTE-DKl … lSG.KSD..IRS ……… VHINWIDSI A0A3L7ZYC0.1 / 38-100 LVT.VS … P-NDDG..SF … VMHLDEK … TDLYPS..NM.ST … SPFG.-K..K … E.VRALV.NFDE —E ….. VA.SSD..IRR ……… VHINWIDSI C9MTD3.1 / 39-104 LVT.IK … TNSSTG..QV … YLQLDDE … TTVLPT..NM.KT … SPYG.N-..K … E.LRALT.NLKIQDG ….. QG.GHY..SKS ……… AYVNWIDTI A0A3A5QVA3.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NA.QG..wSNKD.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVRGINPI D6D3F1.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NA.QG..wSNKD.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVRGINPI A0A0J9FDA3.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NA.QG..wSNKD.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVRGINPI A0A1Y4VS72.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NA.QG..wSNKD.WV..E … G.QRAFV.IFNELEE ….. PV.NGY..DLN ……… IQVRGINPI C3QL57.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NA.QG..wSNKD.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVRGINPI A0A3A5TDG2.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NA.QG..wSNKD.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVRGINPI F7M5N2.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NA.QG..wSNKD.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVRGINPI W4PT47.1 / 59-116 WGT.VK … T-TGGG..GY … YLEGDSW … GTIYPV..AT.SI … PWYR.PV..D … G.QRVIS.FFNPLSD …..—. MKD..KKG …… … VQV —— X5DG99.1 / 30-94 -GT.VMd.yAGTGDC..GF … IIELDNG … NKVQPL..YY.P -… DDFT.FS..Q … G.QRVLI.TYTELNN … ..VY.ISC..DQG ……… VPCEVTY— A0A069SJD8.1 / 37-103 WAT.VN … V-KGAH..VY … DFTGDRW … GQIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD. …. NYpEGY..DCS ……… VKVLSIKEI R9HLR6.1 / 37-103 WAT.VN … V-KGAH..VY … DFTGDRW … GQIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD. …. NYpEGY..DCS ……… VKVLSIKEI F0F989.1 / 39-103 LVT.IK … TNSSTG..QV … YFQLDDE … TTLLPT..NM.KT … SPYG.-N..R … E.VRALT.NIRVQDG ….. QS.GHY..SKS ……… ВЫВНРИДТ- A0A0P0M3E6.1 / 37-103 WAT.VN … V-KGAH..VY … DFTGDRW … GQIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD. …. NYpEGY..DCS ……… VKVLSIKEI A6L4K8.1 / 37-103 WAT.VN … V-KGAH..VY … DFTGDRW … GQIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD. …. NYpEGY..DCS ……… VKVLSIKEI A0A3A5SUL8.1 / 37-103 WAT.VN … V-KGAH..VY … DFTGDRW … GQIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD. …. NYpEGY..DCS ……… VKVLSIKEI A0A1M6H6L2.1 / 39-104 -GT.LE … I-IDGK..DY … FFNLDDG … DKMYPS..DT.AY..iHNYE.LV..N … G.QRVFI.HFLPLEE … ..DK.PGY..EYN ……… ВКЛИКЛЕНИ U2DII9.1 / 59-116 WGT.VK … T-TGGG..GY … YLEGDSW … GTIYPV..AT.SI … PWYR.PV..D … G.QRVIS.FFNPLSD. ….—. MKD..KKG ……… VQV —— A0A3N2MZQ4.1 / 38-103 LVT.VE … P-ADDE..SF … IMHLDDN … TILQPV..NM.AK … SPYG.-K..K … E.VRALV.NFSETVA …. pEN.GGS..FRK ……… VYVNWLDSI C3R4L9.2 / 37-103 WAT.VN … V-KGAH..VY … DFTGDRW … GQIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD. …. NYpEGY..DCS ………VKVLNIKEI I9QI90.1 / 37-103 WAT.VN … V-KGAH..VY … DFTGDRW … GQIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD. …. NYpEGY..DCS ……… VKVLNIKEI B6W4S8.1 / 39-105 WAT.VN … V-KGAH..VY … DFTGDRW … GQIWPA..TT.DY … FWYS.PI..D … G.QRVIL.YFNPLYD. …. NYpEGY..DCS ……… VKVLNIKEI G5SNA9.1 / 42-105 LVT.VK … GGTDS -..- C … FLQLDDE … TTLWPV..NL.SS … SPFG.EK ..-… E.VRALV.HFTDVDG. …. EK.HFC..TRA ……… VYVNWMDSI R6FRW3.1 / 39-102 WAT.VR … T — TGN..AF … YLDCDVW … GTLWPV..NV.DL … GWYE.AV..D … G.QRVIT.VFNPLSD ….. EY.AGY..DHA ……… VKILSMTE- I9S877.1 / 37-99 WAT.VR … V-VSGD..TY … YLESDNY … GSLWPA..AT.NV … YGFK.PV..D … G.KRMLV.VFNPLYD. …. NY.GEY..DMA ……… ИКВЕНЛ — h2HQT2.1 / 44-109 LVT.IK … TNSSTG..QV … YFQLDES … TTILPV..NM.KK … SRYG.-D..K … E.LRALT.NFRVQEG .. … QS.GHY..SKS ……… AYVNWVDTI F2KX91.1 / 39-104 LVT.IK … TNSPTG..QV … YFQLDDE … TTILPT..NM.KT … SPYG.N — .. K … E.VRALT.NIRVQDG .. … QN.GHY..SKS ……… AYVNWVDTI R5MAR8.1 / 39-105 IGT.IK … V-IEDK..EY … YFNLDDG … EKMYPS..DT.TY..iHNYT.VN..D … D.QRVFI.HFLPLEE ….. SI.PGY..KYN ……… VKLIQIEDI A0A415ICF4.1 / 39-105 IGT.IK … V-IEDK..EY … YFNLDDG … EKMYPS..DT.TY..iHNYT.VN..D … D.QRVFI.HFLPLEE .. … SI.PGY..KYN ……… ВКЛИКИЭДИ B3CBU5.1 / 42-108 IGT.IK … V-IEDK..EY … YFNLDDG … EKMYPS..DT.TY..iHNYT.VN..D … D.QRVFI.HFLPLEE .. … SI.PGY..KYN ……… ВКЛИКИЭДИ U6RTN8.1 / 37-99 WAT.VR … V-VSGD..TY … YFESDNY … GSLWPA..AT.NV … YGFK.PV..D … G.KRMLV.VFNPLYD. …. NY.GEY..DMA ……… ИКВЕНЛ — R7DWS9.1 / 65-131 IGT.IK … V-IEDK..EY … YFNLDDG … EKMYPS..DT.TY..iHNYT.VN..D … D.QRVFI.HFLPLEE ….. SI.PGY..KYN ……… VKLIQIEDI A0A1C7H047.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NA.QG..wSNKD.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DFN ……… IQVRGINPI R7J6N1.1 / 38-101 IMT.VV … P — DGS..TY … YLRRDNG … EKLWPF..AT.NC … PGYN.FS ..-… K.TRAQV.NYTMLSD ….. SI.PGF..SHG ……… IKINWIENI A0A6C0RBC1.1 / 31-94 —T.VVd.yAGAGDC..GF … IIELDNG … NKVQPL..YY.P -… DSFT.FS..Q … G.QRVLV.TYTELDN .. … VY.VSC..DQG ……… VPCEVTY— E5VEN4.1 / 44-108 WAT.VR … V — TGN..AF … YLNCDVW … GTLWPV..NT.DI … GWYD.AV..D … G.QRVIT.VFNPLWD ….. NY.AGY..DHA ……. ..VKLLRLQNV A0A1G8L5B1.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVKGINPI B3JPH6.1 / 37-101 WVT.IR … V-EGEG..VY … SFTGDTW … GTMYPA..AN.AV … WNYS.LV..E … G.QRAFL.VFNPLFD. …. НЫ.ЫГЫ..ДВГ ……… ИКПЕРИЙП- R6C6M7.1 / 37-101 WVT.IR … V-EGEG..VY … SFTGDTW … GTMYPA..AN.AV … WNYS.LV..E … G.QRAFL.VFNPLFD. …. НЫ.ЫГЫ..ДВГ ……… ИКПЕРИЙП- A7M622.1 / 44-104 —.—…— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV.IFNELEE ….. PV.NGY .. DLN ……… IQVKGINPI I8YS73.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVKGINPI A0A373GDI9.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVKGINPI A0A373G5Z4.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVKGINPI E5C425.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV.IFNELEE. …. PV.NGY..DLN ……… IQVKGINPI R6JAG6.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVKGINPI A0A1F0IHE9.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVKGINPI A0A3A5SFF5.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. ИФНЕЛИ ….. PV.NGY..DLN ……… IQVKGINPI C3QQ26.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVKGINPI D7K2D2.1 / 44-104 —. — …— QDSK..EF … YFTLDDG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLN ……… IQVKGINPI R6U1h5.1 / 29-88 -VT.IK … PVAQNG.yPF … YMQLDDS … TVIYPS..NL.KQ … SPFG.N — .. K … E.VRAFI.NYKKESE .. …—. EKG..AQV ……… GFINW —- R5ML52.1 / 37-101 WVT.VR … V-EGEG..VY … SFTGDTW … GTMFPA..AN.GI … WNYS.LV..E … G.QRAFL.VFNPLFD….. NY.YGY..DVG ……… IKAEQIYP- A0A4Y8L511.1 / 37-103 LAT.VH..pIDADSK..TY … YLTLDDG … TTLWPA..AS.NV …- IYS.PR..G … D.QRVIV.NYTLLSD … ..QI.GEY..DHY ……… AKINGIQEI A0A412CMU4.1 / 44-104 —. — …— QDSK..EF … YFTLDNG … KKMYPS..NS.QG..wNNED.WV..E … G.QRAFV. IFNELEE ….. PV.NGY..DLS ……… IQVKGINPI A0A078RZE3.1 / 34-99 -GT.VK … V-IEGK..DY … YFSLDEG … SKLYPS..DT.TY..vHDYA.VI..D … G.QRAFI.YFRELEE. …. КЛ.ПГИ..ЭЙН ……… АКИХИЕНИ A0A3N2NFJ2.1 / 38-101 -VT.VV … P-MSDN..GF … ELNLTNT … EKLIPS..NL.KT … SPFK.N-..K … E.VRALV.NFRYDDE ….. TE.KST..SRN ……… VTINWIDSI D3IKU8.1 / 28-100 LVT.VK … PKALDN..SF … VLQLDDS … TQLIPS..KM.KT … SPYG.-S..K … E.VRALV.NYNVVAG .. … PS.AKK..EDGrvd..ertyVNINWLDSI D1QVI7.1 / 38-103 LVT.IK … TNSSTG..QV … YFQLDDS … TTILPV..NI.NK … SLYG.-K..K … E.LRALT.NFKVQEG .. … QS.GHY..SKS ……… AYVNWVDTI A0A174UPB0.1 / 39-104 -GT.VK … V-IEGK..DY … YFSLDEG … SKLYPS..DT.TY..vHDYA.VI..D … G.QRAFI.YFRELEE. …. КЛ.ПГИ..ЭЙН ……… АКИХИЕНИ A0A3A5YD20.1 / 39-104 -GT.VK … V-IEGK..DY … YFSLDEG … SKLYPS..DT.TY..vHDYA.VT..D … G.QRAFI.YFRELEE ….. KL.PGY..EYN ……… AQIKHIENI A7VAA5.1 / 39-104 -GT.VK … V-IEGK..DY … YFSLDEG … SKLYPS..DT.TY..vHDYA.VT..D … G.QRAFI.YFRELEE. …. КЛ.ПГИ..ЭЙН ……… АКИХИЕНИ E5VD66.1 / 39-104 -GT.VK … V-IEGK..DY … YFSLDEG … SKLYPS..DT.TY..vHDYA.VT..D … G.QRAFI.YFRELEE. …. КЛ.ПГИ..ЭЙН ……… АКИХИЕНИ R7EQR9.1 / 39-104 -GT.VK … V-IEGK..DY … YFSLDEG … SKLYPS..DT.TY..vHDYA.VT..D … G.QRAFI.YFRELEE. …. КЛ.ПГИ..ЭЙН ……… АКИХИЕНИ F3PS09.1 / 55-118 WAT.VR … V — TGS..AF … YLDCDVW … GTLWPV..NT.NM … GWYN.PA..D … G.QRVIT.TFNPLWD ….. NY.EGF..DHA ……. ..VKILTLEQ- A0A417LDM2.1 / 42-110 -AT.IK … P-LGGN..TY … YLQWDDS … TTFWPA..AG.AT … PYFG.VD..K … E.RRVFI.NFTLLGDst .sgGI.KGY..DYA ……… IRVNRIDSV E6SP14.1 / 51-109 —. — …—- GN..DY … YFALDEG … SRLYPG..DT.TA..lHNYT.II..D … G. QRAFV.YFDKLNE ….. NK.EGY..EYN ……… AQIKHIENI A0A1Y4JSX4.1 / 38-102 WAT.VR … V — TGN..AF … YLDCDVW … GTLWPV..NT.DL … GWYE.PV..D … G.KRVIT.MFNPLSD ….. GF.DGY..DHA ……… VKLLSLQDV F3ZTI6.1 / 35-103 FGT.VQ … MLDNAD..QGs.lFFLLDSN … QALYPK..NP.GA..vASLD.LQ..D … K.DRVFV.QFTDLEE ….. NH.EKY..DLY …. ….. ANIQIVQK- W4UUV3.1 / 19-81 —. — … PDPKGT..EF … YFGLDDG … KKMYPG..DI.KS..iGDYK.PI..D … G.QRAFV.IFNSFEE. …. KV.AGY..DYN ……… IQVRKIVDI R6LCQ4.1 / 55-119 WAT.VR … V — TGN..AF … YLDCDVW … GTLWPV..NT.DL … GWYE.PV..D … G.KRVIT.MFNPLSD ….. GF.DGY..DHA ……… VKLLSLQDV C9PTP9.1 / 38-110 LVT.VK … PNLGNT..AF … TLQLDDS … VSLTPS..NM.KS … SPFG.MK ..-… E.MRALV.NYNVVDGp … aVK.EGK..DRVkg …. ttyVNINWLDSI D7NEN2.1 / 69-134 LVT.IK … TNSSTG..QV … YFQLDDS … TTILPV..NI.DK … SLYG.-K..K … E.LRALT.NFKVQEG …. .QS.GHY..SKS ……… AYVNWVDTI A0A3A5YF63.1 / 44-108 WAT.VR … V — TGN..AF … YLNCDVW … GTLWPV..NT.DI … GWYD.AV..D … G.QRVIT.MFNPLWD ….. DY.AGY..DHA ……… VKLLRLQNV A7AIT3.1 / 41-102 IVT.VK … P-LSDN..SY … YMQLDDS … TTLFPT..NS.YV … PE-NL — .. K … E.IRAFV.IYKNDEK ….. KT.EGY..DQS ……… VQLLRMD— K5ZE19.1 / 41-102 IVT.VK … P-LSDN..SY … YMQLDDS … TTLFPT..NS.YV … PE-NL — .. K … E.IRAFV.IYKNDEK ….. KT.EGY..DQS ……… VQLLRMD— A0A6G7X532.1 / 38-104 IAT.VN..aIDSTSG..TF … YLTLDNG … STLLPV..AS.D -… SYYK.PQ..F … N.KRVIV.NYTLLSD … ..KT.GVY..DHA ……… IKINAIQDI R7EJU1.1 / 38-102 WAT.VR … V — TGN..AF … YLNCDVW … GTLWPV..NT.DI … GWYD.AV..D … G.QRVIT.VFNPLWD ….. DY.AGY..DHA ……… VKLLRLQNV A0A174EQQ7.1 / 38-102 WAT.VR … V — TGN..AF … YLNCDVW … GTLWPV..NT.DI … GWYD.AV..D … G.QRVIT.VFNPLWD ….. DY.AGY..DHA ……… VKLLRLQNV A0A069DAR1.1 / 39-104 MAT.VK … V-INGA..NY … YLVLDNN … KTVWPG..AT.AV … PQYQ.AV..D … G.QRVIA.NITLLGD….. QY.GSY..DYV ……… AKINYLNNV R5IQH6.1 / 40-104 -GN.VV … S-EEGK..PS … YLLLDSK … RTMLFS..EN.SI … PSLY.LE..D … G.RRVIV.NYTLLDE ….. DY.QGY..DFR ……… IHVNDIDTV A7V7U3.1 / 44-108 WAT.VR … V — TGN..AF … YLNCDVW … GTLWPV..NT.DI … GWYD.AV..D … G.QRVIT.VFNPLWD ….. DY.AGY..DHA ……… VKLLRLQNV A0A380ZBT7.1 / 30-94 WAT.VR … V — TGN..AF … YLDCDVW … GTLWPV..NT.DL … GWYE.PV..D … G.KRVIT.IFNPLSD ….. GF.DGY..DHA ……… VKLLRLYDV A0A2V1IPK5.1 / 42-104 LVT.VC … P-TVNG..GF … TMVLDDK … TTLVPS..NM.DK … SPFG.N-..K … E.VRALV.CYTPDKS …..- F.HGG..T-G ……… VTVNRIDSI R6XDY4.1 / 41-102 IVT.VK … P-LSDN..SY … YMQLDDS … TTLFPT..NS.YV … PE-NL — .. K … E.IRAFV.VYKNDEK ….. KT.EGY..DQS ……… VQLLRMD— R7LG97.1 / 77-140 -VT.VC … PTPNG — .. SY … FMQLDDS … TTLLPK..GN.PP … APYG.-K..K … D.VRALV.NYEEAYG ….. GR.RGG..RQT ……… VNVLWMDSI E5WXU5.1 / 38-102 WAT.VR … V — TGN..AF … YLDCDVW … GTLWPV..NT.DL … GWYE.PV..D … G.KRVIT.IFNPLSD ….. GF.DGY..DHA ……… VKLLRLYDV A0A4R2E341.1 / 35-101 -GT.IQ..kVSPNAQ..PF … FILLDNG … DKVWPA..VT.NF … PTYN.PK..E … G.QRIIA.GYSILTN ….. GS.NDY..NHD ……… IKLYDVYEV A0A1M6B9L8.1 / 40-98 -AT.VR … L — TGN..AF … YLNSDTW … GTLWPI..NT.DL … GWYQ.PV..D … G.QRMVT. ИФНПИЙД ….. НЙ.ГЕЙ..ДХА ……… ВКИЛ —— A0A3R5ZK82.1 / 41-110 IAT.IK … P-LGND..AY … YLQWDDS … TTFWPA..SG.VT … PYFG.VD..K … E.RRAFI.NFTLLGDst. шМА.ЕГИ..ДЯ ……… ВРВНРИДСВ S0GLY8.1 / 41-110 IAT.IK … P-LGND..AY … YLQWDDS … TTFWPA..SG.VT … PYFG.VD..K … E.RRAFI.NFTLLGDst. шМА.ЕГИ..ДЯ ……… ВРВНРИДСВ R5IKT4.1 / 34-102 IVT.VKpieNPDDSL..TF … YMQLDDS … TTLVAE..NL.KK … SPFG.-D..K … E.VRALT.SIEETEA ….. TV.PGY..DKV …… … ВХВЯЙДСИ E6STJ1.1 / 39-102 -AT.MR … V — TGS..SF … YLDCDVW … GTLWPV..NT.DM … GWYE.PA..D … G.QRVIT. VFNPLWD ….. NY.EGF..DHA ……… VKILRLQNV A0A1Y4I2B4.1 / 38-103 LAT.VR … A-VSGT..GF … YLDCDVW … GTCWPV..NT.DM … GWYI.PI..D … G.QRVVT.VFNPLWD. …. NY.GGF..DHA ……… VKILEIQNV U2ING5.1 / 41-106 LVT.IK … TNSSTG..QV … YFQLNDE … TTILPT..NM.KT … SPYG.-K..K … E.LRALT.NIKVQDG .. … KT.EGY..SKS ……… AYVNWVDTI W4P9B5.1 / 43-105 —.—… PDAEKK..EF … YLTCDNG … ETLYPY..NV.GR..lGNYK.LV..D … G.QRAFV.ACSFVDE ….. KV.EGY..DHT. …….. ВЕЙФЕЙВДИ G1WE61.1 / 41-106 LVT.IK … TNTATG..QV … YLQLDDA … TTVLPT..NM.KT … SRYG.-N..R … E.LRALA.NLTMQEG .. … QP.GHY..SKT ……… AFVNWMDTI A0A069SN58.1 / 39-101 MAT.VE … KQPQYD..AP … YLVLDNG … EKLWVV..QH.IV … PYRD.LK..A … G.ERIFG.NYSFLEA … ..GE.SGF..AYN ……… IRLND —- A0A5D3FVX9.1 / 42-104 —. — … PDAEKK..EF … YLTCDNG … ETLYPY..NV.GR..lGNYK.LV..D … G.QRAFV.ACSFVDE. …. КВ.ЕГИ..ДХТ ……… ВЕЙФЕЙВДИ A0A374UXL0.1 / 38-101 WAT.VR … V — TGN..AF … YLDCDAW … GTLWPV..NT.DL … GWYE.PV..D … G.KRVIT.VFNPLSD .. … GF.DGY..DHA ……… VKLLRLHD- R5RT16.1 / 38-98 -VT.VK … PEGEGS..GF … YLQLDDN … TIIKPV..NN.PR … LPFE.LK ..-… E.TRAFI.NYKVVSE .. …—. EKG..VQM ……… GHVNWIT— A0A0P0M594.1 / 44-116 LAT.VV … TPSEIP..GE..aMIESDND … GVAYVV..NP.DKl.tRFET.NN..P … G.QRIFY.TYINADN …..- P.TGD..ASKkg ….. pfISIDYLQKI A0A3A5SVY8.1 / 44-116 LAT.VV … TPSEIP..GE..aMIESDND … GVAYVV..NP.DKl.tRFET.NN..P … G.QRIFY.TYINADN …..- P.TGD..ASKkg …..pfISIDYLQKI R9HIS3.1 / 44-116 LAT.VV … TPSEIP..GE..aMIESDND … GVAYVV..NP.DKl.tRFET.NN..P … G.QRIFY.TYINADN …..- P.TGD..ASKkg ….. pfISIDYLQKI U6REE9.1 / 35-101 IAT.LK … A-TDAD..SY … YFGLDSG … RKMYPG..DD.SA..vRNYP.IV..D … G.KRVFV.WFHQLEE .. … QV.PGY..DYN ……… IQVVQMDSI A6KWC3.1 / 44-116 LAT.VV … TPSEIP..GE..aMIESDND … GVAYVV..NP.DKl.tRFET.NN..P … G.QRIFY.TYINADN …..- P.TGD..ASKkg ….. pfISIDYLQKI I8XQ39.1 / 35-101 IAT.LK … A-TDAD..SY … YFGLDSG … RKMYPG..DD.SA..vRNYP.IV..D … G.KRVFV.WFHQLEE….. QV.PGY..DYN ……… IQVVQMDSI A0A2N8N7W0.1 / 26-91 -AT.VD … PVGRGG..TY … SLTLDDG … SRLWPA..GT.EI … PFYR.PN..E … R.HRALA.YYTILSD .. … TF.QGF..DHA ……… ILVRSIRDI A0A0N7J6Z6.1 / 39-101 MAT.VE … KQPQYD..AP … YLVLDNG … EKLWVV..QH.IV … PYRD.LK..A … G.ERIFG.NYSFLEA … ..GE.SGF..AYN ……… IRLND —- R9HHX7.1 / 39-101 MAT.VE … KQPQYD..AP … YLVLDNG … EKLWVV..QH.IV … PYRD.LK..A … G.ERIFG.NYSFLEA … ..GE.SGF..AYN ……… IRLND —- A6L1M8.1 / 39-101 MAT.VE … KQPQYD..AP … YLVLDNG … EKLWVV..QH.IV … PYRD.LK..A … G.ERIFG.NYSFLEA ….. GE.SGF..AYN ……… IRLND —- A0A180F7F0.1 / 38-104 IVT.IE … SLNEGA..DF … FYRLDDG … STLWPS..AG.YY … RGHN.LE..D … G.QRALL.NYTILSD … ..SL.DGF..SHF ……… ВКВНСИДПИ K6A328.1 / 41-102 IVT.VK … P-LSDN..SY … YMQLDDS … TTLFPT..NS.Y -… MPEN.LK ..-… E.IRAFV.IYKNDEK ….. KT.EGY..DQS ……… VQLLRMD— A0A1G4GB27.1 / 35-96 IAT.VI … P-TGES..GY … RLLLDNG … KQLWPV..AT.DV …- YYL.PS..D … D.QRVFV.NYTVLSG ….. RQ.GNY..DHY ……… ВКВНДИ — A0A3A5SNJ1.1 / 39-101 MAT.VE … KQPQYD..AP … YLVLDNG … EKLWVV..QH.IV … PYRD.LK..A … G.ERIFG.NYSFLEA ….. GE.SGF..AYN ……… IRLND —- A0A3P2A1D1.1 / 48-114 IGT.VK … V-IEGK..EY … YFALDEG … SKLYPG..DT.TY..iHNYV.IV..D … G.QRAFV.NFSKLSE .. … TI.KGY..HYN ……… GKIHRIENI A0A078SU12.1 / 23-87 WAT.VR … V — TGN..AF … YLNCDVW … GTLWPV..NT.DM … GWYD.AV..D … G.QRVIT.VFNPLWD ….. DY.AGY..DHA ……… VKLLRLQNV A0A562VGU0.1 / 45-111 IGT.VR … V-IEGK..EY … YFALDEG … SKLYPG..DT.TS..iHNYA.IV..D … G.QRAFV.NFSKLNE .. … TI.KGY..HYN ……… GKIHRIENI A0A239RKH7.1 / 41-107 -VT.LK … PQPSGL.. — … LLQLDDS … TTLWAV..NM.KQ … SPYG.-S..K … E.LRAFV.NYREPKEse.qvPQ.QGT..AKN ……… VFVNWIDTI A0A4S2B5P2.1 / 38-103 -GT.VK … V-IEGK..DY … YFSLDEG … SKLYPS..DT.TY..vHNYA.VI..D … G.QRTFI.YFHELEE. …. КЛ.ПГИ..ЭЙН ……… АКИХИЕНИ A0A318HTY5.1 / 38-110 LVT.VK … PGTGTT..PF … ILQLNDS … TSLMPY..NM.RT … SPFG.-D..K … E.VRALV.NYDVMYS .. …—. VGQ..RSGgapvkdatfVTINWIDSI A0A1H5VRG8.1 / 39-103 -AT.VV … P-INDN..SF … YLRTDDG … STLWPA..AP.IY … IHYE.AK..E … R.QRAWV.NYTILGD ….. СИ.ГНИ..ШС ……… ИКИНГИСДИ E2NHZ5.1 / 39-104-GT.IE … V-IGEK..EY … FFNLDDG … DKMYPS..DT.TY..iHNYT.VE..N … N.QRVFI.HFLPLEE ….. DI.PGY..DYN ……… ВКЛИКЛЕНИ F3ZSP5.1 / 39-102 YAT.VV … R — EGN..TY … SLKLDSG … QKLWVG..AG.WI …- NYN.SV..D … G.QRVVA. DYTILGD ….. NY.NGY..DHI ……… VKVNYLYEI I9FQV4.1 / 39-104 -GT.IE … V-IGEK..EY … FFNLDDG … DKMYPS..DT.TY..iHNYT.VE..N … N.QRVFI.HFLPLEE. …. DI.PGY..DYN ……… ВКЛИКЛЕНИ R6KMN2.1 / 39-104 -GT.IE … V-IGEK..EY … FFNLDDG … DKMYPS..DT.TY..iHNYT.VE..N … N.QRVFI.HFLPLEE. …. DI.PGY..DYN ……… ВКЛИКЛЕНИ A0A0D8J6J7.1 / 13-71 —. — … AGAGDC..GF … IIELDNG … RKVQPL..YY.P -… DDFT.FS..Q … G.QRVLI.SYTELNN. …. VY.ISC..DQG ……… VPCEVTY— W2CM13.1 / 40-105 -VT.VD … PIGKGG..TY … SLTLDDG … TRLWPA..GT.EI … PFYR.PT..T … R.HRALA.YYTILSD .. … TF.QGY..DHA ……… ILMRSIRDI A0A015WFR3.1 / 42-112 IGT.MK … I-PNAD..TPrdfFIALDNG … DNVLPA..DT.ADirnRKYT.VA..E … G.QRVFV.GYLQMGE ….. EK.PGY. .EN ……… ГКИФТИЕДИ A0A015XFW5.1 / 42-112 IGT.MK … I-PNAD..TPrdfFIALDNG … DNVLPA..DT.ADirnRKYT.VA..E … G.QRVFV.GYLQMGE ….. EK.PGY. .EN ………ГКИФТИЕДИ W0EST4.1 / 36-103 IVT.CH … RGNDGT..YF … THQATDSaplDTLYPV..G-.AV … NEAK.TA..E … G.VRVLL.QYRPVET …..- L.GEN..RQR ……… IEIQALSSI U6RJD8.1 / 37-101 WAT.VE … A-KDSH..TY … SLTGDTW … GTLWPA..AS.AF … PW-S.PE..D … G.ERVVI. VFNPLYD ….. DF.QGY..DCA ……… IKIEGIRSI A0A373IXR6.1 / 37-101 WAT.VE … A-KDSH..TY … SLTGDTW … GTLWPA..AS.AF … PW-S.PE..D … G.ERVVI. VFNPLYD ….. DF.QGY..DCA ……… IKIEGIRSI R6XKL5.1 / 41-103 LVT.IK … T-TDSG..VT … YFQLDKK … TTLEPT..NW.K -… NPYK .— .. R … E.VRALL. QYSEEPG….. EV.GLF..SKK ……… VKVAWIDSV A0A5D3E7Q2.1 / 37-94 WAT.VK … T-TGGG..GY … YLEGDSW … GIIYPV..AT.SI … PWYR.PV..N … G.QRVIS.FFNPLGD. ….—. MKD..KKG ……… VQV —— W4P6L2.1 / 59-116 WAT.VK … T-TGGG..GY … YLEGDSW … GIIYPV..AT.SI … PWYR.PV..N … G.QRVIS.FFNPLGD. ….—. MKD..KKG ……… VQV —— A0A1R3T9X3.1 / 36-101 IAT.VV … P-EGEN..NY … SFLLDNG … KRLWPA..AS.NI …- RYT.PK..D … N.QRVFL.NYTILWD ….. SPgTTY..DYD ……… IKVNDIWNI A0A180EYW2.1 / 38-104 IVT.IE … SLNDGA..DL … FYRLDDG … STLWPS..AG.YY … RGHN.LE..D … G.QRALL.NYTILSD ….. SL.DGF..SHF ……… VKVNSIDPI A0A017NFB3.1 / 42-112 IGT.MK … I-PNAD..TPrdfFIALDNG … DNVLPA..DT.ADirnRKYT.VA..E … G.QRVFV.GYLQMGK ….. EK.PGY. .EN ……… ГКИФТИЕДИ Q5LFQ7.1 / 42-112 IGT.MK … I-PNAD..TPrdfFIALDNG … DNVLPA..DT.ADirnRKYT.VA..E … G.QRVFV.GYLQMGK ….. EK.PGY. .EN ……… ГКИФТИЕДИ A0A3R6RK43.1 / 41-110 IAT.IK … PLGNNN ..- Y … YLQWDDS … TTLWPA..SG.VT … PDFG.VD..K … E.RRAFI.NFTLLGDsi.ssGI .PGY..DYI ……… IRVNKIDSV C3REN2.1 / 39-101 MAT.VE … KQPQYD..AP … YFILDNG … EKLWVV..QN.AV … PYRD.LK..T … G.ERIFG.SYTFLEA ….. GE.SGF..AYD ……… IRLND —- I9Q8F6.1 / 39-101 MAT.VE … KQPQYD..AP … YFILDNG … EKLWVV..QN.AV … PYRD.LK..T … G.ERIFG.SYTFLEA … ..GE.SGF..AYD ……… IRLND —- W2C6Y9.1 / 40-97 -VT.VD … PIGKGG..TY … SLTLDDG … TRLWPA..GT.EI … PFYR.PT..T … R.HRALA.YYTILSD .. … TF.QGY..DHA ……… Я ——— B6W0T7.1 / 32-94 MAT.VE … KQPQYD..AP … YFILDNG … EKLWVV..QN.AV … PYRD.LK..T … G.ERIFG.SYTFLEA … ..GE.SGF..VYD ……… IRLND —- A0A3L7Z5E7.1 / 37-89 WAT.VH … T-TGGG..GY … YLEGDRW … GTIDPV..ST.SI … PWYK.PV..D … G.ERVVA.FFNPLYD ….. VE.NG -.. — …. …..——— A0A1Y4HPS0.1 / 52-111 —. — …— EGN..DY … YFELDEG … SKLYPG..DT.TA..iSNYP.LV..E … N.QRAFV .YFTLLEQ ….. AL.PGY..EYN ……… ААЛХВЕНИ R5KEE0.1 / 38-98 FVE.AQ … TDNNAN..IV … SIETDDG … ERMFLT.gAI.NV..aWAER.PD..T … V.YRALL.YYNKVEA ….. -D.DGT..NNA ……… EP ——- A0A264Y8K6.1 / 51-115 LVT.VK … PDNSGM..RF … MMQLDDS … TTINAV..NI.QK … APYD .— .. K … E.VRAFV.SFRMLTP .. … PD.KRR..HYD ……… VYVNWMDSI A0A180EY81.1 / 42-108 IVT.IE … SLNDGA..DL … FYRLDDG … STLWPS..AG.YY … RGHN.LE..D … G.QRALL.NYTILSD ….. SL.DGF..SHF. …….. ВКВНСИДЛИ A0A374VBB3.1 / 39-103 WAT.VR … V — TGN..AF … YLDCDVW … GKLWPT..NV.NL … GGYE.PA..D … G.QRVLT.VFNPLVK ….. ЭЙ.ГДФ..ДЯ ……… ВНИЛРЛКЕВ W4UTP9.1 / 34-93 WAT.VQ … A-TGGG..EY … TLVGDKW … GTILPV..AT.AI … PWYK.PV..D … G.ERVVA.FFRPLEE. …. GA.QGQ..EHD ……… VVV —— W2CFD6.1 / 40-97 -VT.VD … PIGKGG..TY … SLTLDDG … TRLWPA..GT.EI … PFYR.PT..T … R.HRALA.YYTILSD .. … TF.QGY..DHA ……… Я ——— F5IUY9.1 / 35-99 IVT.VK … K-IDDN..TY … SFVRDNG … DKLWVA..AP.VG … LNLK.PK ..-… Y.DRAII.NYTILSD … ..EK.DGY..DHY ……… IRLNDYYDV A0A1h3QC26.1 / 36-99 WAT.VH … A-TGGG..GY … YLEGDRW … GVIDPV..AS.SI … PWYK.PV..D … G.ERVVA.FFNPLAD. …. TD.NGA ..— Q ……… ВКИЕГИКЕВ A0A174W3Q0.1 / 35-98 FVT.VK … AGTDGS..IQ … TLIPDNG … EALTVS..KD.RT..gSAIS.PN..T … S.RRVMS.NYETLSN …. . — .- GH..TAT ……… AVIYSLQS- G5GAU4.1 / 39-104 LVT.IK … TNSSTG..QV … YFQLDDS … TTVLPT..NS.IN … LSY-.VK..K … E.LRALA.NLKMQNG .. … QS.APY..SKS ……… АЙВНВИДТИ A0A4Q0TW54.1 / 42-113 IGT.MK … I-PNAD..TPrdfFIALDNG … NNVFPA..DT.ADirnWKYT.VA..E … G.QRVFV.GYVKMEG …. eEK.PGY .. EN ……… ГКИФТИЕДИ R5RXV8.1 / 42-113 IGT.MK … I-PNAD..TPrdfFIALDNG … NNVFPA..DT.ADirnWKYT.VA..E … G.QRVFV.GYVKMEG …. eEK.PGY .. EN ……… ГКИФТИЕДИ E4VQB0.1 / 42-113 IGT.MK … I-PNAD..TPrdfFIALDNG … NNVFPA..DT.ADirnWKYT.VA..E … G.QRVFV.GYVKMEG …. eEK.PGY .. EN ……… ГКИФТИЕДИ R6DDQ8.1 / 36-88 WAT.VH … A-TGGG..GY … YLKGDRW … GTIDPV..ST.SI … PWYK.PV..D … G.ERVVA.FFNPLYD….. ВЕ.ГГ -.. — ………——— E4T748.1 / 31-99 IAT.VE … NPTQST..AF … DFRLDDR … TFMH-T..NT.AG..lNYK.PK..D … G.QRIIA.NYSFIEKv .. .sPT.SNS..VNV ……… VKLNDVYEI R7CW41.1 / 34-99 MAT.VT … D-RDG — .. RW … VLESDNC … GELIPL..NT.AA..lTMHDaDK..D … N.QRVIA.AFNYVDE … ..- К.ДРС..НКГ ……… ИЕЛЫДЫЙРВ A0A1Y4D9M9.1 / 40-101 —T.VV … P — EGN..TY … YLLRDNG … ETLWPF..AS.NE … MDYV.PS ..-… H.TRASV .NYTLLSD ….. SI.PGF..SHG ……… IKVNWIENI A0A6H0KM28.1 / 37-89 WAT.VH … T-TGGG..GY … YLEGDRW … GTIDPV..ST.SI … PWYK.PV..D … G.ERVVA.FFNPLVD ….. VE.GG -.. — ………——— A0A143XJM8.1 / 33-102 FAT.VA..vPAQSSR..NY … YFVLDDG … ATVYPS..DK.DRi.mYNAN.GK..N … G.NRAVI.YYNFLAS ….. SV .PGF..DHN ……… IALYDVVDI A0A5R9R915.1 / 37-102 -GT.VH … K — TAD..YY … YITTDGG … SNLWPS..AS.NV … NSED.LE..D … G.MRVLV. НИТИЛГДа … дДС.ЭФЫ..ДЫЫ ……… ВКВНГВСЕИ E4M9L1.1 / 40-98 —. — …—- SN..DY … YFVLDDD … KTVYPS..DK.SR..vAGYK.PE..D … D. QRVII.FFNLLKT ….. GV.EGY..DYN ……… IALYDVRNI A0A553EHX2.1 / 36-92 WAT.VH … T-TGGG..GY … YLEGDRW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD ….. ME.GG -..- KG ……… VQ —— — A0A1Y4AHI9.1 / 38-101 WVT.VH … V-QSDG..VY … TFTGDTW … GTMFPA..AP.TV … WGYR.LA..E … G.TRGLL.VFNPLYD. …. DY.YGY..DVG ……… IKPQQLR— R5V7L7.1 / 36-98 —.- E … TL-NSN..DY … YFVLDDN … KTVYPS..DK.SR..vAGYK.PE..D … D.QRVII. FFNLLKT ….. GV.EGY..DYN ……… IALYDARNI A0A1Y3VD89.1 / 37-104 —T.VC … PLDGGD ..- Y … CFQRDNG … DKLYPS.dKT.RV … PGYR.PA..DdgtK.QRAII.WFSLLNE ….. TV.PGY..RYN ……… IALYAVEEI F0R3T6.1 / 38-101 WVT.VH … V-QSDG..VY … TFTGDTW … GTMFPA..AS.VV … WGYR.LS..E … G.TRGLL.VFNPLYD ….. NY.YGY..DVG ……. ..IKPQQLR— R5BZ69.1 / 38-107 LVT.VK … HDAEG — .. VF … YLQLDDK … TTAKPS..NL.KK … SPFG.EK ..-… Q.VRALA.HLTVEDQpstdiNN.SRF ..DKV ……… VMVDWIDSV B6VTS4.1 / 44-116 LAT.VV … TPSEVP..GE..aVIESDND … GIAYVV..NP.DKl.tRFET.NN..P … G.QRIFY.TYINAAN …..- P.TGD..ASKkg ….. pfISIDYLQKI C3R8A1.1 / 44-116 LAT.VV … TPSEVP..GE..aVIESDND … GIAYVV..NP.DKl.tRFET.NN..P … G.QRIFY.TYINAAN …..- P.TGD..ASKkg ….. pfISIDYLQKI I9QWT0.1 / 44-116 LAT.VV … TPSEVP..GE..aVIESDND … GIAYVV..NP.DKl.tRFET.NN..P … G.QRIFY.TYINAAN …..- P.TGD..ASKkg … ..pfISIDYLQKI A0A069SJL2.1 / 44-116 LAT.VV … TPSEVP..GE..aVIESDND … GIAYVV..NP.DKl.tRFET.NN..P … G.QRIFY.TYINAAN …..- P.TGD..ASKkg ….. pfISIDYLQKI A0A374W6V7.1 / 52-114 MAT.VA … K-ETGT..VY..pYFVLDNG … KTVWVA..AS.AV … EYEG.LE..P … G.QRVIG.NFTILDD .. … NR.DGF..DYF ……… ARVNN —- A0A401LYR5.1 / 37-89 WAT.VH … A-TGGG..GF … YLEGDRW … GIIDPV..TT.AI … PWYR.PV..D … G.KRVVA.IFNPLAD. …. TK.DG -.. — ………——— A0A0L8V6A4.1 / 39-105 IVD.VE … T — GSD ..- Y … TIKMDDG … SELFPV..NS.AI … HWHE.ED..D … S.ARVLV.NYTILDEk. .rvND.EFE..EYY ……… VRVNSVKDI C3QHJ8.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- D6D137.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- F7M3D3.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ………VQ ——- A0A0J9FK80.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- A0A1h5FR43.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- A0A373GGU0.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- A0A3A5QFD2.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD….. ME.GG -..- KG ……… VQ ——- A0A3A5TGE9.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- C3QZP6.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- A0A1C7GY13.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- A0A1G6G9U1.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD ….. ME.GG -..- KG ……… VQ ——- R6JM30.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDNW … GVIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- F8X179.1 / 39-98 MVT.VN … K-VGEN..TY … DFTLDNG … KKLWIA..AP.VG … LNLK.PK ..-… Y.NRAII.NYTILSD. …. QY.QGY..DHA ……… IKLN —— A0A1M4UDU9.1 / 35-98 FVT.VK … SGSDGK..LQ … TLLPDNG … DEMLVS..KD.RT..nSTIS.PN..N … S.KRVLA.NYEVLSH …. .—. NGV..AS ……… ATIYSLQS- A0A1M5BYQ7.1 / 39-101 LGT.VK … V-DAGN ..- V … SFVLDNG … EKLWPA..AT.LV … PYKN.LE..N … G.TRIIG.NFTLLSD ….. AQ.NGF..DHY ……… VRLND-YS- R6YT86.1 / 36-105 -PT.LG … TVEDVD..PI … TIASDTY … GTLIPT..NP.EV … FEKS.GV..Ds.iG.QRVFL.NVVFQSDp … hNP .IST..AQE ……… VIVLNLYNI A0A401LV94.1 / 35-98 FVT.VK … SGTDGK..LQ … TLLPDKG … DEMLVS..KD.RT..nSTIS.PN..N … S.KRVLA.NYEVLSS …. .—. NGV..AS ……… ATIYSLQS- A0A1Y4UAF9.1 / 36-102 —T.VC … PLEGGD ..- Y … CFERDNG … DRLYPS..DK.SR..vPGYK.PDgdQ … K.QRAII.WFSLLDQ …. .TI.PGY..RYN ……… IALYAVEEI A0A1M4UZP1.1 / 37-88 ВАТ.VH … A-TGGG..GF … YLEGDRW … GIIDPV..TT.AI … PWYR.PV..D … G.KRVVA.VFNPLVD ….. TK.D —.. — … ……——— Q8A0L6.1 / 36-98 WAT.VR … A-TGGG..GY … YLEGDRW … GMIDPV..AS.SI … PWYK.PV..D … G.ERVVA.FFNPLAD. …. TD.KGA ..— Q ……… VKIEGIQE- A0A1T5HSS5.1 / 34-98 -GT.IE … K-GDIS..PY … VVVADNG … DRLFPS..AT.AV … PHFK.TK..D … G.QRVWI.SYTILGD ….. AE.GQI..DHY ……… VRVNDLSEI A0A373ZYJ3.1 / 35-100 —T.VR … TLDGGD ..- Y … YFQRDNG … ETLYPG..NK.SR..vPGYK.PDpdK … T.QRVII.WFTLQNG …. .-I.ADY..DYN ……… IDLYFVENI D4IN28.1 / 38-103 —Т.VR … TLDGGD ..- Y … YFQRDNG … ETLYPG..NK.SR..vPGYK.PDpdK … T.QRVII.WFTLQNG …..- I.ADY..DYN … …… ИДЛИФВЕНИ A0A4Y1X463.1 / 35-97 -TT.VH … T-LGDK..EY … YFERDNG … QTLYPS..EK.S -… ATFE.AE..E … G.KRAVI. YFDLLEG …..- I.PDY..DYN ……… IRLYRAEEI R5JFL8.1 / 37-99 WAT.VR … V-LSGN..TY … YLDSDYY … GTLWPA..AS.NV … YGFR.PV..D … G.QRLAV.LFNPLYD. …. НФ.ДНЫ..ДЛА ……… ВКВЕНВ — A0A1Y3WL00.1 / 37-104 IVT.CH … HHDGNT..YF … THQATDSapmDTLYPT..ET.PN … ESLT.-A..E … G.VRVLL.QYKPLES …..- L.GEN..RER ……… IEIQALSAI A0A3M8EDM0.1 / 36-98 IAT.VV … P-EGEN..AY … SFLLDNG … KRLWPA..AG.NV … RYAP.-T..R … N.RRVFL.NYTILWE .. … ССГАНЫ..ДЫД ……… ИКВНДВ — A0A1h4ZCB3.1 / 44-105 —. — …- GETGT..DY … YFQRNNG … EKLYPG..DK.TR..iPNYK.GK..E … R.QRAII.WFKLLEE ….. RV.PDY..DYN ……… ЯЛЯВДЕЙ A0A139L7W9.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG …. . — .— .. ЛОР ……… АТИЙСЛК— R6KCB5.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG …. . — .— .. ЛОР ………ATIYSLQ— A0A3A5S8G9.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG …. . — .— .. ЛОР ……… АТИЙСЛК— A0A373GKQ6.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG …. . — .— .. ЛОР ……… АТИЙСЛК— A0A1F0I3S5.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG …. . — .— .. ЛОР ……… АТИЙСЛК— A7M0A4.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG….. — .— .. ЛОР ……… АТИЙСЛК— Д7К4х2.1 / 35-95 ФВТ.ВЕ … АГЕДГР..IQ … ТЛИПДКГ … ЭВЛТВС..ЭД.РТ..гСТИС.ПН..Т … С.РРВМС.НЕВЛСГ …. . — .— .. ЛОР ……… АТИЙСЛК— E5C919.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG …. . — .— .. ЛОР ……… АТИЙСЛК— I9SSG9.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG …. . — .— .. ЛОР ……… АТИЙСЛК— A0A373GC20.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG ….. — .— .. ENT ……… ATIYSLQ— A0A412CRA8.1 / 35-95 FVT.VE … AGEDGR..IQ … TLIPDKG … EVLTVS..ED.RT..gSTIS.PN..T … S.RRVMS.NYEVLSG …. . — .— .. ЛОР ……… АТИЙСЛК— R5PV50.1 / 42-100 —. VE … AFTSSDtiVM … KAVTDDG … ASLTFT..QR.IK..tSWTSvPD..S … S.YRALL.YYSKGDN …..— — P..VRA ……… LSLR —— A0A1Y4h4B0.1 / 36-103 IVT.CH … RGNEGT..YF … TCQATDSspmDTLCPI..G-.AV … DESR.TA..E … G.VRVLL.QYRPVEE …..- L.STT..RKR ……… IEVQALSAI W7YLW6.1 / 39-103 —T.VL … M — DVD..SY … SIKTDNG … KVLWPS..AS.NV … SVSL.LE..D … S.MRVLV.NYTILGEa … tDN.DSY..DYY ……… VRVNGLSKI A0A412CZ32.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDRW … GLIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- E5C6T9.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDRW … GLIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- A0A1Y4AM38.1 / 51-116 -AT.VH … PLGEN — .. NY … YFTTDSN … RSIYAG..DK.SR..iIAYE.PV..E … G.QRAFI.WFNYLPV .. … AF.EGY..DYN ……… VELYKIENI A0A1I4D1U4.1 / 36-101 IAT.VI … P-EGEN..AY … SFLLDND … KRLWPA..AG.NV … RYA-.PT..H … N.QRVFL.NYTILWE ….. SSgATY..DYD …….. .IKVNDIWNI A0A1F0IXY7.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDRW … GLIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- A0A373FY59.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDRW … GLIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- A7M3Y4.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDRW … GLIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- D7K892.1 / 36-92 ВАТ.VR … A-TGGG..GY … YLEGDRW … GLIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD ….. ME.GG-. .-Кг ……… VQ ——- I9T0S7.1 / 36-92 WAT.VR … A-TGGG..GY … YLEGDRW … GLIDPV..AT.SI … PWFK.PV..D … G.ERVVA.FFNPLYD. …. ME.GG -..- KG ……… VQ ——- R5JCG5.1 / 36-102 -AT.LR … QIDDTD..DY … YFGLDSG … RKMYPG..DD.SA..iRNYP.IV..D … G.KRVFV.WFHQFEE … ..QV.PGY..DYN ……… IQVVRMDSI R5UPD5.1 / 57-117 —. — …— KNGE..LT … TIQTDRG … ALVCPS..VW.NI …- PAE.FK..N … K. DRVLI.YFNLLGEa … dST.EQF..DYY ……… VRINEIHKI A0A0E9LSV0.1 / 37-101 -AT.ID … T-EEVE..PF … LIITDNG … DRLFPS..AT.AV … PGFG.YD..H … G.QRVWV.SYTILGD .. … GE.GDV..DHL ……… VKVNDMVEI A0A174T2G8.1 / 37-94 WAT.VR … T-TGGG..GY … YLVGDRW … GIIDPV..AT.SI … PWFK.PV..D … G.ERVVS.FFNPLAD. ….—. MSD..EKG ……… VQV —— A0A6F9Z7N3.1 / 40-112 LAT.VV … TPSEVP..GE..aVIESDND … GIAYVM..NP.DKl.tRLET.NN..P … G.QRIFY.TYTNAAN …..- P.IGD..ASKkg ….. pfISIDYLQKI K5C9h4.1 / 38-95 WAT.VR … T-TGGG..GY … FLEGDRW … GIIDPV..SS.SI … PWYR.PV..D … G.ERVVA.FFNPLAD. ….—. МКД..ЭКГ ………VQV —— A0A174AYE5.1 / 38-95 WAT.VR … T-TGGG..GY … FLEGDRW … GIIDPV..SS.SI … PWYR.PV..D … G.ERVVA.FFNPLAD. ….—. MKD..EKG ……… VQV —— C9KSP9.1 / 38-95 WAT.VR … T-TGGG..GY … FLEGDRW … GIIDPV..SS.SI … PWYR.PV..D … G.ERVVA.FFNPLAD. ….—. МКД..ЭКГ ……… VQV —— A0A1Y3X3N5.1 / 38-100 WVT.IH … V-ESDG..IY … TFTGDTW … GTMYPA..AS.GI … WGYQ.IT..E … G.TRGLL.VFNPLYD. …. NY.YGY..DVG ……… IKPEQL — A0A1Y4FLU8.1 / 38-100 WVT.IH … V-ESDG..IY … TFTGDTW … GTMYPA..AS.GI … WGYQ.IT..E … G.TRGLL.VFNPLYD….. NY.YGY..DVG ……… IKPEQL — A0A1Y4D741.1 / 38-100 WVT.IH … V-ESDG..IY … TFTGDTW … GTMYPA..AS.GI … WGYQ.IT..E … G.TRGLL.VFNPLYD. …. NY.YGY..DVG ……… IKPEQL — R5URh3.1 / 38-100 WVT.IH … V-ESDG..IY … TFTGDTW … GTMYPA..AS.GI … WGYQ.IT..E … G.TRGLL.VFNPLYD. …. NY.YGY..DVG ……… IKPEQL — A0A6I6JSQ7.1 / 41-107 —. — … TESNAN..GY … KIVMDND … DLLIPV..TS.NF..nVFHY.VE..D … G.DRVLV.NYTILDDns. geGE.EAT..EYY ……… ИКЛНСВККИ A0A0F5JKU2.1 / 42-110 -AT.VK … P-LGNN..SF … YLQWDDS … TSFWPA..AG.VT..tPDFD.VS..K … E.RRAFI.NFTLLGPg..kdMN.IEQ..DYV ……… IRLNRIDTV X5E6B2.1 / 39-105 —. — … KGDDAG..AV … SLVMDNG … SVLIPA..AA.TRp.gWYLD.FT..D … G.DRVLV.NYTILDE .. … DE.SSS..AVEr …… ylVKVNDISDV R6ZZW4.1 / 34-98 FID.VT … TNADGT..LQ … NLITDKG … EALQIL..NR.DG..lDGLT.PD..S … T.YRTVS.IYEPKEM …. .NA.QGN..AT ……… ALLYSCQ— R6SCC0.1 / 31-97 -ID.AR … TNGEGK..LT … WLITDQG … DTLDIQ..ER.SG..lDGLC.AD..S … T.YRTIT.IYEHLEE … ..EA.SQT..YPM ……… VKVYSVQQI S0F6X7.1 / 31-97 -ID.AR … TNGEGK..LT … WLITDQG … DTLDIQ..ER.SG..lDGLC.AD..S … T.YRTIT.IYEHLEE ….. EA.SQT..YPM ……… VKVYSVQQI A0A521BDQ3.1 / 41-101 —. — …— EVG..SF … YVTTDEG … KKLWPS..AT.NV … NVSL.LE..D … G. MRVLV.NYTILGDa … nLD.SAY..DYL ……… VKINDVSQI A0A6H0KKX8.1 / 35-94 FVT.VV … AGEDGS..IQ … TLIPDKG … ASLPVL..ED.RT..gAAIS.PN..T … S.RRVMS.NYEVLTD …. . -. — ..— G ……… VRIYSLQS- A0A1h3Y0V5.1 / 36-98 FVT.VK … AGADGL..IK … SLLPDKG … KLLTVS..KD.RT..gSSIS.PN..T … A.RRVIS.NYEVNSD …. . — .— .. ДАТ ……… AVIYSLQSV Q8A1F1.1 / 36-98 FVT.VK … AGADGL..IQ … SLLPDKG … ELLTVA..KD.RT..gSTIS.PN..S … A.RRVIS.NYEVNPE ….. — .— .. DAT ….. …. AVIYSLQSV R6ZNW4.1 / 34-97 LAE.AQ … TNDKGT..LT … YLLTDKG … DKYRVL..ER.EG..lEGLT.PD..S … I.YRALC.VYQVTDT …. -. QG -..- KT ……… VQLYSAQKV A0A015WI52.1 / 25-86 -LT.AE … SGADGR..LQ … TLVTDKG … ERLVVA..ED.RT..eSELL.PN..S … S.SRVVS.NYEVLSS … ..—. ВГГ..РКЕ ……… ИРИЯЛ — A0A3L7Z2Y1.1 / 35-94 FVT.VV … SGEDGS..IQ … TLIPDKG … ESLPVL..ED.RT..kSAIS.PN..T … S.RRVMS.NYELLTG …. . -. — ..— G ……… VRIYSLQS- Q5LI29.1 / 37-98 -LT.AE … SGADGR..LQ … TLVTDKG … ERLVVA..ED.RT..eSELL.PN..S … S.SRVVS.NYEVLSS …..—. VGG..RKE. …….. ИРИЯЛ — A0A017NJW6.1 / 37-98 -LT.AE … SGADGR..LQ … TLVTDKG … ERLVVA..ED.RT..gSELL.PN..S … S.SRVVS.NYEVLSS … ..—. ВГГ..РКЕ ……… ИРИЯЛ — A0A015UC55.1 / 37-98 -LT.AE … SGADGR..LQ … TLVTDKG … ERLVVA..ED.RT..eSELL.PN..S … S.SRVVS.NYEVLSS … ..—. ВГГ..РКЕ ……… ИРИЯЛ — R6MGN3.1 / 36-99 LGT.VN … A — DGK..DI … IIESDSY … GPLVPV..NP.GFi.tSTQT.DS..T … G.QRIMA.VFQYAGEt .. .aEK.TSN..STS ……… ВЕЛИ —— R6D3S3.1 / 35-94 FVT.VV … SGEDGS..IQ … TLIPDKG … ESLPVL..ED.RT..kSAIS.PN..T … S.RRVMS.NYELLTG …..- -. — ..— G ……… VRIYSLQS- U2JHB1.1 / 33-85 FVE.AK … TNELSA..FT … SAITDDG … TQLTLK..PA.VKe.qWATK.PD..T … T.YRALL.YYNKVEN ….. Г-.—..—………——— A0A2P8CCI4.1 / 47-110 —. — …- NPSDN..SY … VIQLDNG … DSFVPM..AT.SV … PYFN.VY..N … N.QRVWV. NFNPLDDk … sNS.DGS.lTYY ……… GKVNDIREI A0A255T0T4.1 / 38-91 FVE.AH … TNDDTM ..- F..dYVITDNG … DSLRLT..IPvET..sWAKK.PD..S … L.YRAML.YYNKVEG ….. NK . — .. — ………——— B3JJR7.1 / 36-88 LGT.VN … A-DEND ..- I … TIESDSY … GLLIPT..NP.GFi.tSTQT.DS..T … G.QRILA.VFQYTEN .. … ТТ.К —.. — ………——— R6CFN9.1 / 36-88 LGT.VN … A-DEND ..- I … TIESDSY … GLLIPT..NP.GFi.tSTQT.DS..T … G.QRILA.VFQYTEN .. … ТТ.К —.. — ………——— A0A4Q0TTM0.1 / 36-98 FLT.AE … AGADGT..LQ … TLVTDKG … ERLVVA..ED.RT..rTELF.PN..G … S.SRVVS.NYEVISS …. .—. AGG..QKE ……… VRIYAL — W4UVJ3.1 / 35-98 FVT.VE … AGADGT..LQ … RLIPDKG … EALPIS..KD.RT..gSTIA.SN..S … S.RRVLS.NYEKVSS….. — .- GG..VSS ……… AVIYAVQS- E4VXM8.1 / 36-98 FLT.AE … AGADGT..LQ … TLVTDKG … ERLVVA..ED.RT..rTELF.PN..G … S.SRVVS.NYEVISS …. .—. AGG..QKE ……… ВРИЯЛ — R5RVR4.1 / 36-98 FLT.AE … AGADGT..LQ … TLVTDKG … ERLVVA..ED.RT..rTELF.PN..G … S.SRVVS.NYEVISS …. .—. AGG..QKE ……… VRIYAL — A0A4Q8RM95.1 / 38-101 —. IE … G — DQD..SY … IIVTDSG … DRLFPS..AS.NV … PNYP.VE..D … D.MRVWV .NYTILGDa … tDN.ET.lDYY ……… VKINDLSDV A0A399SVR7.1 / 31-92 -GT.VMd.yAGTGGC..GI … VIELDNG … TVIQPL..YY.PK..gFIFT .—..E … G.QRVLI.EYSKLSK ….. VV.NKC..GEG ……… ISCE —— A0A255TTF4.1 / 37-90 FVE.AH … VNSEKK..ID … YAITDNG … DSLVIS..TPiEV..sWTKT.AD..S … T.YRAIL.YYNKMAN ….. NT . — .. — ………——— A0A096AX82.1 / 33-85 FVE.AK … TNELSA..FT … SAITDDG … TQLTLK..PA.VKe.qWAKR.PD..T … T.YRALL.YYNKVEN ….. Г-.—..—………——— A0A174IN52.1 / 35-94 FVT.IE … AGDDGR..IR … TLIPDKG … DILPVS..QD.LT..gSTIS.PN..T … S.RRVMS.NYETMEE …. . -. — ..— G ……… AKIYSLQS- C9KZ00.1 / 35-94 FVT.IE … AGDDGR..IR … TLIPDKG … DILPVS..QD.LT..gSTIS.PN..T … S.RRVMS.NYETMEE ….. -. — ..— G ……… AKIYSLQS- A0A255T6J3.1 / 38-103 -VT.VK … PNADNS..SF … VMQLNDS … VQIMAS..NM.VT … SPFG.TK ..-… E.KRALI.GFTAADG .. ..qQT.EGE..LTR ……… VKVLWMDSI A0A6C0RE73.1 / 40-105 —. — …- GDGEG..VG … SLVMDNG … YVLVPA..AT.SGs.gWFHN.IS..D … G.DRILV.NYTILDE. …. DE.TSS..SEEr …… yfVKINDISDV A0A3E4MT54.1 / 35-98 -ID.AA … TDESGT..LQ … KLITDKG … ESLQIL..NR.EG..lDGLT.PD..S … I.YRSVS.IYEPKES … ..AD.EGQ..AT ……… ALLYSCQ— R5VVP4.1 / 36-98 —N.AA … TDESGT..LQ … KLITDKG … ESLQIL..NR.EG..lDGLT.PD..S … I.YRSVS.IYEPKES ….. AD.EGQ..AT …….. .ALLYSCQ— R9I7H6.1 / 44-116 LAT.VV … NPSEVP..GE..aVIESDND … GIAYVV..NP.DKl.tRLET.NN..L … G.QRIFY.TYINAAN …..- P.TGD..ANNkg ….. pfISIDYLQKV A0A553ES39.1 / 35-95 FVT.VK … AGDDGR..IQ … TLIPDKG … EALSVA..ED.RT..gSTIA.AN..T … S.RRVMS.NYEVLSG …. . — .- G -..- SA ……… ATIYSLQ— A0A5M4AKK8.1 / 47-110 —. — …- SPDVN..SY … VIQLDNG … DSFVPM..AT.SV … PYFN.VY..D … N.QRVWV. NFNPLDDk … tNS.DGS.lTYY ……… GKVNDVREI K5DC43.1 / 35-94 FVT.IE … AGDDGR..IR … TLIPDKG … DILPVS..QD.LT..gSTIS.PN..T … S.RRVMS.NYETMEE ….. — .— .. —G ……… AKIYSLQS- A0A0D8J5G3.1 / 39-105 —. — … NGDDAG..AV … SLVMDNG … SVLIPV..AA.SSp.gWSSH.FS..D … G.DRVLV.NYTILDD .. … ДД.ЦС..АВЕР …… ЫлВКВНДИСДВ A0A4U2L0K8.1 / 44-108 —. — …- DGDAQ..LF … EIELDND … DRLIPV..AG.HL … HYINrVE..D … N.DRVLV.NYTILDD. …. AD.TDN..DPSt …… fyVRINDIDKV R5B9R0.1 / 42-104 LVT.IK … T-AESG..ET … YFQVNEK … TTLSPV..NW.T -… NPYK .— .. R … E.VRALL. RYSEERG ….. DG.GLF..SKK ……… VKVAWIDSV R6EX48.1 / 45-93 -VL.LH … TSQDKS..VR … SALLDDG … TTLQFS..NPfAQ … DWLTrPD..S … V.YRALL.YYDKT —…..- -. — .. — ………——— A0A1M4VFF5.1 / 37-87 —. — …- GMNYC..GF … VIELDNG … TKIQPH..NY.PK … DFE-.FV..Q … G.QRILV .DYVMLPN ….. II.STC..DKG ………——— R5F5E6.1 / 36-103 IVT.CH … TNKGNP..YF … TYQTTDSapvDTLYPV..S-.EV … NSDD.MG..E … G.VRVLL.QYRPIET …..- L.SEH..KKQ ……… VEIQALSAI A0A0K8QSP7.1 / 43-103 —. — …—- PN..TF … TVLMDDG … SVIYPV..AG.HYn.gDWFD .— .. D … D. SRVLV.NYTIVDDk..kvDD.TNK..EYY ……… ВКИНСВРКИ A0A0J9IDV0.1 / 35-96 FVT.VE … AGEDGR..IQ … TLIPDKG … EALPVA..ED.RT..gSTIA.AN..T … S.RRVMS.NYEVLPD …. . — .- G -..- SA ……… ATIYSLQS- A0A174HA24.1 / 35-96 FVT.VE … AGEDGR..IQ … TLIPDKG … EALPVA..ED.RT..gSTIA.AN..T … S.RRVMS.NYEVLPD …. . — .- G -..- SA ……… ATIYSLQS- C3QY43.1 / 37-98 FVT.VE … AGEDGR..IQ … TLIPDKG … EALPVA..ED.RT..gSTIA.AN..T … S.RRVMS.NYEVLPD …. . — .- G -..- SA ……… ATIYSLQS- A0A3A5TQL0.1 / 35-96 FVT.VE … AGEDGR..IQ … TLIPDKG … EALPVA..ED.RT..gSTIA.AN..T … S.RRVMS.NYEVLPD….. — .- G -..- SA ……… ATIYSLQS- C3QEh4.2 / 35-96 FVT.VE … AGEDGR..IQ … TLIPDKG … EALPVA..ED.RT..gSTIA.AN..T … S.RRVMS.NYEVLPD …. . — .- G -..- SA ……… ATIYSLQS- D6D6C0.1 / 35-96 FVT.VE … AGEDGR..IQ … TLIPDKG … EALPVA..ED.RT..gSTIA.AN..T … S.RRVMS.NYEVLPD …. . — .- G -..- SA ……… ATIYSLQS- F7MAI2.1 / 35-96 FVT.VE … AGEDGR..IQ … TLIPDKG … EALPVA..ED.RT..gSTIA.AN..T … S.RRVMS.NYEVLPD …. . — .- G -..- SA ……… ATIYSLQS- A0A3A5QD27.1 / 35-96 FVT.VE … AGEDGR..IQ … TLIPDKG … EALPVA..ED.RT..gSTIA.AN..T … S.RRVMS.NYEVLPD ….. — .- G -..- SA ……… ATIYSLQS- R6MKT1.1 / 36-98 —. AQ … TDSDGT..IS … QLVADNG … TVYPVL..QR.DG..lGGLV.AD..T … L.YRTIS.IYEPLPQ .. …- E.DGK..ETA ……… AQLYSCQL- A0A1I2FQ17.1 / 39-104 —. IN … A-ESGS..GY … TIKMDDG … SELFPV..AG.YP … YWDD.LE..D … S.TRVLV. NFTILDDk..tvDE.EHE..QYY ……… VRVNSVKDI G6AJT2.1 / 46-104 FAE.VS … TNASSA..FV … EAVTDDG … TTLRLTpaYR.PK … WKVK.AD..T … T.YRALL.YYNKVED …..- -.- GA..TAV ……… VA ——- A0A3M8EG83.1 / 34-86 FAT.VV … K-ENSN ..- Y … RFRLDNG … RLLIPE..A -.- V … KDYS.GK..E … G.QRVIL.NYTPLEG ….. -. — ..- DA ……… IKINF— — G6B0S0.1 / 47-95 -VL.LH … TSQDKS..VH … SALLDDG … TTLQFS..NPfAQ … DWLTrPD..S … V.YRALL.YYDKT —…. . -. — .. — ………——— R5CUU2.1 / 31-88 FVE.AE … TDASAK..VV … SIVTDRD … ERLWLT..RS.VKl.dWAEK.PD..T … A.YRSLI.YYNKVKD ….. GN.GQY..V —………——— R5MYK7.1 / 36-98 LGT.VN … A — DGK..DI … KIESDSY … GLLIPI..NP.GFi.tSTQT.DS..T … G.QRVLT.VFQYAGSa .. .tEN.ESN..STS ……… VEL —— Э7РЛГ9.1 / 35-90 ФВЕ.АХ … Т-СТВГ..VF..dYVVTDDG … DTLSMQ..PYmKV..sWAQT.PD..S … L.YRCLL.YYNHVNG …..- Y.AEP .. — …….. .——— A0A6I6JVP4.1 / 31-87 —T.VVd.yAGAASC..GI … VIELDNG … NRIQPL..YY.P -… DGFT.FT..Q … G.QRVLV.EYKELPN .. … VI.PTC..DRG ………——— A0A069SDB7.1 / 33-97 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTD … ..- S.EEA..EKE ……… AMLYNTQL- A0A3A5SGP9.1 / 33-97 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTD … ..- S.EEA..EKE ……… AMLYNTQL- A6KXU5.1 / 33-97 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTD …..- S.EEA..EKE. …….. AMLYNTQL- C3REX0.1 / 33-97 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTD … ..- S.EEA..EKE ……… AMLYNTQL- B5D3W3.1 / 35-98 -ID.AA … TDENGT..LQ … KLITDKG … ESLEIL..NR.EG..lDGLT.PD..S … I.YRSVS.IYEPKES … ..AD.KGQ..AT ……… ALLYSCQ— R5FN37.1 / 54-115 LAM.VH … TMDRQI..AD … YMLTDEK … DSVHFS..PL.AR..lDYAE.RA..D … SfYRSLV.YYNKVSS …..- -.- КА..ССА ……… FAIQQ —- A0A1G4G915.1 / 34-87 -AT.VI … Q — TAD..RL … TFQLDNF … KTLVPI..DP.R -…- GYS.GK..D … G.QRVIL. NYSPLRG ….. -. — ..- DT ……… IQINSVN— A0A0P0M6I3.1 / 45-109 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTD … ..- S.EEA..EKE ……… AMLYNTQL- R9HA98.1 / 45-109 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTD … ..- S.EEA..EKE ……… AMLYNTQL- A0A6F9Z8K4.1 / 35-99 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTA … ..- P.EET..EKE ………AMLYNTQL- A0A3E4QJB0.1 / 27-78 FVE.AT … TNAAAT..FV … MAETDDG … EKIVLA..PPlKT..kWATS.PD..S … T.YRALL.YYNRFSD …..- -. — .. — ………——— I8VNS6.1 / 33-97 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTD … ..- S.EEA..EKE ……… AILYNTQL- B6VVA8.1 / 33-97 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTD … ..- S.EEA..EKE ……… AILYNTQL- A0A415GP05.1 / 47-95 —. LR … TAPDKS..VR … SVQLDDG … TELRLS..NPfAK..dWIER.PD..T … V.YRALL.YYDQMSA….. -. — .. — ………——— A0A1C7GY40.1 / 35-96 FVT.VK … TGDDGR..IQ … TLIPDKG … EALPVA..ED.RT..gSVIA.AN..T … Y.KRVMS.NYEVLSD …. . — .- G -..- SA ……… ATIYSLQS- A0A399CY63.1 / 40-93 —. — …— SKHC..SF … IIELDNG … EQIQPF..HY.P -… DEFT.FT..Q … G. QRVLI.RYVKLPN ….. II.PTC..DQG ……… VACD —— A0A1M6JRD5.1 / 38-88 —. — …- GAGNC..SF … VIELDNG … NRIQPL..HS.P -… DDFD.FA..H … G.QRVLV .EYSELEN ….. VI.PAC..DRG ………——— A0A3P1SUG3.1 / 40-88 —. TR … TNAAGE..LF … CAVTDNG … DSLVLS..PK.RTa.kWATT.PD..S … I.YRALL.YYNVPEK ….. -. — .. — ………——— A0A1h5CJE8.1 / 36-86 -AE.LA … VNSQQQ..GV … SFVTDEG … EQLALA..NP.LT..aSWITrPD..T … T.YRTIV.YYNKVEN ….. —. — .. — ………——— A0A1H8F3R6.1 / 42-102 FVE.AR … TDANKQ..VV … SMTTDDG … DLLPLV.kPY.SA..kWITT.AD..T … I.YRCML.YYNKVEQ ….. -H.QG — .. QYA ……… AEV —— A0A095ZI76.1 / 37-94 MAD.MH … TAAKCK..VD … YAVTDTD … EKLTFS..PPlNC..hWATT.PD..S … T.YRAML.YYKKSPD ….. KT .EGN..T —………——— R5LSA3.1 / 31-87 FVE.AE … TDASSN..VV … SIMTDRD … ERLWLT..RS.VKl.eWAEK.PD..T … A.YRSLI.YYNKVKD …..- D.NGT..Y —………——— — R9ICA5.1 / 35-99 -ID.LK … TDHTGT..GR … YLITDEG … TEWRIQ..SR.TG..lDGLA.PD..T … T.YRTVT.MYAPLTG … ..- P.EET..EKE ……… AMLYNTQL- A0A0C3RE89.1 / 41-104 —.- H … K-SAED..SY … TVKLDNG … SVIYPS..YS.DV … PTQN.LQ..D … S.TRLLV .DFTILQDa … iPG.SKF..DYY ……… ВРИНГЕРИ I4Z7Q2.1 / 44-87 —. — …—- AQ..AF..aSAITDDN … TSLTLS..PA.IAa.kWATT.PD..S … T.YRAML. ЙЫНКВЕН ….. -. — .. — ………——— R5PDQ9.1 / 65-127 —. — … N-KEGS..SY … TISTDLG … AVLFPS..AS.NV … SSNY.LE..D … K.DRVIV.TFSILGDa … dTS.SMY..DYY ……… ВКВВДВЫКВ E1KPF7.1 / 27-78 FVE.AT … TNAAAT..FV … MAETDDG … EKIVLA..PPlKT..kWATS.PD..S … I.YRALL.YYNRFSD …..- -. — .. — ………——— D1W947.1 / 37-94 MAD.MH … TAAKCK..ID … YAVTDTD … EKLTFS..PPfSC..hWATT.PD..S … T.YRAML.YYKKSPD ….. KT .EGN..T —………——— A0A1I3ZGJ4.1 / 49-101 FAT.VV … K-ENSN ..- Y … RFRLDNG … RLLIPE..E -.- V … KNYS.GE..E … G.QRVIL .NYIPLEG ….. -. — ..- DA ……… IKINF —- F2KUA9.1 / 46-100 FVE.AR … TNTATA..FF … SAHTDDG … VRLTLQ.pAL.KT..lWATK.GD..T … T.YRALL.YYNKVEN ….. GV.T —..— -………——— U2KRh2.1 / 37-89 -AE.LH … TTSPRV..VD … YLITDDG … DSVAFS..PR.ITv.dWAIR.GD..S … V.YRVLA.YYNLSEE …. .GP. — .. — ………——— A0A099BWX5.1 / 67-130 -VT.VK … PMKDGK..KF … YIWMSED … KVGHAK..NY.AK … SPFG.-D..K … E.VRALA.NLREEGK. ….- D.KNG..VLQ ……… FYVNWMDSI G5H8U8.1 / 41-97 —. — …— TQD..AY … TIKTDGG … NTLTVI..EN.LV … PSFP.VV..D … G. MRVRV.NYTIEKQ ….. TA.NGF ..— E ……… VRLNAIEKI R5MZ28.1 / 34-98 -IG.VQ … TDHEGT..IS … QLVADNG … TVYPVL..QR.DG..lGGLV.AD..T … L.YRTVS.IYEPIVQ ….. -E.EGK..ETA ……… AQLYSCQL- A0A1R3T558.1 / 8-59 FAT.VV … K-ENSN ..- Y … RFRLDNG … RLLIPD..EV. — … KDYS.GE..E … G.QRVIL. НИИПЛГГ ….. -. — ..- ДА ……… ИКИН —— A0A2K9HA32.1 / 33-84 FVD.AR … TNELGA..FV … SAKTDDG … EELTLS..PA.VKe.qWATR.PD..T … T.YRALL.YYDRVEK ….. —. — .. — ………——— A0A1I5HBU5.1 / 48-98 -VD.LS … INSEKE..AT … AFTLDDG … NRFTLT..PY.ATa.kWIET.AD..T … T.YRAVL.YYNQVSE …. . -. — .. — ………——— F0R6Y8.1 / 46-101 LGT.VM … M — DNQ..GI … TIESDLY … GNLIPE..NP.ET..iSSIDaDS..T … G.QRILA.GFIFLNN … ..PT.DQS..S —………——— F8NBW9.1 / 44-103 LVT.FK … TDSTTG..AF … FMQLNDS … VTLVDK..NI.TK … SPYG.-D..K … E.VRGLI.SFVPVDK .. … NE.TGL..RRQ ……… МГП —— A0A099C0R1.1 / 35-88 IVD.AH … AATKGQ..ID … YCHTDKG … DLLKLS..PH.AK..aDWATtPD..S … T.YRALL.YYGKVTD ….. HT . — .. — ………——— D9RSP6.1 / 34-85 -VE.AR … TNELSA..FA … SATTDDGi.qLTLKPA..VK.EQ … WAKR.PD..T … T.YRALL.YYNKVEN…..Г-.—..—………——— D5EV56.1 / 43-95 FAE.AV … VNSNQQ..VT … KIVTDDG … DELSLT..AP.YTa.kWVSK.AD..T … T.YRCML.YYNKVDN ….. Q -. — .. — ………——— A0A318HU86.1 / 51-103 FGE.VH … TTAKGT..LA … YVVTDEG … DTLRIA..DN.IV..nKSLAvAD..S … V.YRVLY.YYDQNGK ….. N -. — .. — ………——— A0A249T0A3.1 / 36-77 —. — …— DGC..KW … VIELNDG … TRLQPV..EL.AD … STFK.LS..N … E. QRVKV.SYTELKD ….. -. — .. — ………——— A0A3D8HAN6.1 / 41-102 IVT.VK … PLTDN — .. TY … YMQLNDS … ITLFPT..NS.YV … PE-N.L-..К … Э.ИРАЙВ.ИЛРЕСДК ….. В.В.СДЫ..ДКА ……… ВЕВИРМД— A0A0E9LVC1.1 / 35-99 —. VV … EGNSAS..TF … MIFTDGG … ERLRPV..EF.QP … TNYE.IE..E … G.KRVLA.RYSVIEE. … sDS.ETY..DYS ……… ВКЛАСИСНВ A0A396M211.1 / 46-97 —. — … —— ..- F … KIETDNG … NTLRVT..EN.LD … PSFP.VE..D .. .S.LRVVA.TFTPLEQ …..- T.GEN..SFD ……… IRVNAMK— F9Z8N4.1 / 40-100 —. VS … K-SGDE..KF … TVKLDDG … YLLYPR..EY.YF … NPEK.LN..D … S.TRLRI. RFNILEE …..—. QDT..NLY ……… ARIVYADTI R7GWN5.1 / 47-95 —. LR … TAPDKS..VR … SVQLDDG … SELRLS..NPfAK..dWIER.PD..T … V.YRALL.YYDQMSA ….. -. — .. — ………——- — A0A416A3S4.1 / 41-102 IVT.VK … PLTDN — .. TY … YMQLNDS … ITLFPT..NS.YV..pQNL -.— .. K … E.IRAYV.ILKESDK. …. В.В.СДЫ..ДКА ……… ВЕВИРМД— A0A1Y4AT81.1 / 34-97 FID.IQ … TDESGT..IT … HLVADNG … KRYAVQ..PR.EG..lNGLH.AD..T … I.YRTVS.IYQFLTE …. .KT.NSS..EAE ……… VMLYS —- F0R8T9.1 / 34-98 FID.IQ … TDGSGT..IT … QLAADNG … RHYRVQ..PR.EG..lDGLR.AD..T … I.YRTVS.IYQFLTE …. .KA.SQT..EAE ……… VMLYS-S— R5C6Y9.1 / 40-100 -LT.AY … SGGDGA..LE … RVLTDDG … EEYAVL..VD.DS..hSHIR.PD..S … L.VRIVS.NYEWVHE …..- E.GG — .. EG …… … ВХЛЯВ — A0A1Y4F071.1 / 34-97 FID.IQ … TDESGT..IT … HLVADNG … KRYAVQ..PR.EG..lNGLH.AD..T … I.YRTVS.IYQFLTE …. .KT.NSS..EAE ……… VMLYS —- A0A255TRI3.1 / 43-102 -VT.LK … PTAIGN.hEW … FIQLSDS … SIVTCS..NI.KK … FPYD.NK..K … E.YRAYI.YFKFDSE … .. — .— П..СЛА ……… АТИНСИ — R5I855.1 / 36-96 MVT.CL … I — NNN..SV … YFQLDNG … SYLIPD..NT.IT …- PES.IK..S … M.QRVML. НЫВИКЕК ….. РИ.ТДК..ЭЙИ ……… ИКВРСИ — R7DJE5.1 / 34-97 FVT.VL … S-PEEG..IR … FLRDDGI … VLLSET..TN.DT … DNKY.LV..T … G.KRILL.RYIPGEQ …..- I.SQT. .ELS ……… AKIQSITF- F0F4J1.1 / 46-98 FVE.AR … T-NSAS..AF..iSARTDDG … AQLTLQ..PAlKT … PWATkGD..T … V.YRALL.HYNKVEK ….. G -. — .. — ………——— D1W0R4.1 / 39-90 —E.IH … TSTKNK..VD … YAWTDEG … KKLTFT..VP.FT..cSWAHvPD..S … V.YRAML.YYTQDEE …. .ST. — .. — ………——— A0A1Y4D9E8.1 / 34-96 -ID.LQ … TDATGT..IT … HLIADDG … EHYAVQ..PR.EG..lDGLR.TD..T … L.YRTVS.VYQILNE … ..QT.DGN..EPE ……… AILYS —- L1NKU8.1 / 37-84 FAE.AH … TDANKA..LT … YMVNDAG … DTLRFT..QR.RQ..lKSLNvAD..T … A.YRALV.YYN —-…. . -. — .. — ………——— A0A1Y3XKI3.1 / 34-96 -ID.LQ … TDATGT..IT … HLIADDG … EHYAVQ..PR.EG..lDGLR.AD..T … L.YRTVS.VYQILNE … ..QT.DGN..EPE ……… AILYS —- A0A2T5BSY2.1 / 36-101 FGL.VQ … AGENGE..DF … RIAVDPG … YAIIPI..NL.HE..iNLEK.YK..N … D.VRVIV.NYTIVGDr … tVG .EES..EYY ……… ARINL —- E6SUE3.1 / 36-96 -LT.AY … AGADGS..LQ … SIRTDEG … KAYMVV..ED.AA..kFSMK.AD..S … S.ARIVS.NYEPMVA … ..- A.DGT..SG ……… ВКЛЯ —- R5U564.1 / 34-96 -ID.LQ … TDATGT..IT … HLIADDG … EHYAVQ..PR.EG..lDGLR.AD..T … L.YRTVS.VYQILNE … ..QT.DGN..EPE ……… AILYS —- B3JP22.1 / 36-98 —. AQ … TNNDGT..IS … QLVADDG … TIYSVL..QR.DD..lGGLV.AD..T … L.YRTIS.IYEPITQ .. …- E.NGK..GNV ……… AQLYSCQL- R6CZ20.1 / 36-98 —. AQ … TNNDGT..IS … QLVADDG … TIYSVL..QR.DD..lGGLV.AD..T … L.YRTIS.IYEPITQ .. …- E.NGK..GNV ……… AQLYSCQL- C9PTP6.1 / 51-102 FAE.AH … TATNAT..LA … FAVTDEG … DTLRFP.pNT.SN … KALTkAD..S … V.YRVLY.YYDQRGA….. -. — .. — ………——— S8FCH0.1 / 36-89 LAE.MR … TDKEGN..AQ … TLITDRG … QEWTIR..NE.KK..iGGLV.PD..S … L.YRVLA.RYAQAGE …. . — .- GT..S —………——— A0A3E4N1W4.1 / 47-109 —. — …— NTE..RI … TIDSDTY … GTLLPT..NP.GI … FTEK.KV..Ni.lG.QRVLL. EILFLNE ….. DK.QNNtgDKE ……… VKIISLYKV A0A1M5G1R1.1 / 37-97 —. AQ … TNASGK..II … SLTTDKD … ATYTVE..KD.RT..dSKLS.AN..A … L.QRIVC.NYIATEP .. … AD.NSQ..NAS ……… AVIYS —- A0A1T5D8Y2.1 / 44-102 —. — …—— S..TF … LIEADDG … TILRPI..EY.PS … DYDE.II..E … G.KRVFF.YFKHVDAg … rDD.QNF..DYS ……… IRIQAMQDV I4Z7T2.1 / 40-103 -VT.IK … PINNGE..GF … NIWVSDK … VQGQAT..NL.VK … SPYG.N — .. K … E.VRALA.NIRDEGK. ….- A.ANG..MQK ……… IYVNWLDSI F8N725.1 / 40-88 —.- K … TDHQAR..IT … SAITDDG … TSLTFT..QPlQV..nWKLT.PN..S … L.CRTML.YYNMYAN … ..—.-Г-..—………——— A0A098BZB4.1 / 34-83 FAT.VI … K — QGD..TY … RFKLDNG … KVLSPI..EV.K -… DFHG.-S..D … G.DRVIL .YWTPLNG ….. D -. — ..— K ……… IE ——- U6RBJ9.1 / 33-94 -ID.LV … TDNTGT..AT … YLKSDEG … VIWSIQ..PR.AG..lDGLT.PD..S … V.YRTVT.MYAPVNT ….. SQ.P-S..SLE ……… AILYN —- A0A373IK38.1 / 33-94 -ID.LV … TDNTGT..AT … YLKSDEG … VIWSIQ..PR.AG..lDGLT.PD..S … V.YRTVT.MYAPVNT … ..SQ.PS..SLE ……… AILYN —- R5AXI1.1 / 56-100 —. — … —— .. — …—- DNG … TTLLTA..SR.AL … DTAT.VD. .P … G.DRILA.YYTPEEP …..- D.IRL..ADY ……… APVNT —- B5D292.1 / 42-110 —. LG … TVVNTE..QL … TIDSDTY … GALLPV..NT.HIf.tDNKV.NI..L … G.QRVLL.EVLFLDK … ..NK.QNN..NSEa …… kkVEIISLYKV U2P836.1 / 44-94 -VE.AA … TNGNSQ..FV … QALTDDN … ARLAFT..KP.VKa.aWATT.PD..S … T.YRALL.YHKQVAA ….. -. — .. — …. …..——— //
Как В-клетки формируют иммунный ответ против Mycobacterium tuberculosis
Eur J Immunol. Авторская рукопись; доступно в PMC 2009 12 октября.
Опубликован в окончательной редакции как:
PMCID: PMC2760469
NIHMSID: NIHMS137197
Кафедра микробиологии и иммунологии Медицинского колледжа Альберта Эйнштейна
Отделение для переписки: Медицина, микробиология и иммунология, Медицинский колледж Альберта Эйнштейна, Форххаймер 406, 1300 Morris Park Ave, Bronx, NY, 10461
Окончательная отредактированная версия этой статьи издателем доступна бесплатно в Eur J Immunol. См. Другие статьи в PMC, в которых цитируется опубликованная статья .Abstract
Обширная работа, демонстрирующая важность клеточных иммунных механизмов для защиты от Mycobacterium tuberculosis , в значительной степени отодвинула биологию В-клеток на второй план в области борьбы с туберкулезом (ТБ). Однако недавние исследования показали, что В-лимфоциты посредством различных взаимодействий с клеточным иммунным ответом играют ранее недооцененную роль в формировании защиты хозяина от невирусных внутриклеточных патогенов, включая M.Туберкулез . Работа в нашей лаборатории недавно показала, что, рассматривая эти лимфоциты в более широком смысле в рамках их разнообразных взаимодействий с клеточным иммунитетом, В-клетки оказывают значительное влияние на исход заражения вирусом M. tuberculosis , а также на результирующий воспалительный ответ. В этом обзоре мы выступаем за пересмотренный взгляд на иммунологию туберкулеза, в котором роли клеточного и гуморального иммунитета не исключают друг друга. В контексте нашего текущего понимания защиты хозяина от невирусных внутриклеточных инфекций, мы рассматриваем недавние данные, подтверждающие более значительную роль В-клеток во время M.tuberculosis , чем считалось ранее.
Ключевые слова: туберкулез, микобактерии, В-клетки, гуморальные, рецепторы Fcγ
Введение
Несмотря на жизненно важное значение для иммунологической защиты и вакцинации против широкого спектра микробных патогенов, В-клетки остаются загадкой в области туберкулеза. Традиционно считается, что В-клетки и антитела не вносят значительного вклада в защиту от M. tuberculosis [1]. Несмотря на ранние доказательства того, что гуморальный иммунитет может влиять на микобактериальную инфекцию, противоречивые результаты пассивной иммунной терапии (вероятно, из-за вариабельности подготовки антисывороток), проведенной в конце XIX века [2], привели к его замене более безопасным и более устойчивым антибиотиком. лечение туберкулеза к 1950-м годам [3].Со временем усилия основоположников иммунологии, таких как Альберт Кальметт и Эли Мечников, подняли клеточный иммунитет на передний план защиты хозяина в двадцатом веке [4-6], где с тех пор он доминирует в исследованиях в области туберкулеза [7]. Недавний рост молекулярной генетики лишь еще больше снизил значение В-клеток для борьбы с туберкулезом, поскольку исследования иммунодефицита В-клеток у людей [8, 9] и мышей [10, 11] поставили под вопрос, оказывают ли эти лимфоциты защитный эффект. против М.Туберкулез .
Успехи иммунологии в двадцать первом веке привели исследователей к рассмотрению не только естественного иммунитета к инфекции, который сам по себе может варьироваться с точки зрения защиты [12], но и новых способов рационального улучшения стратегий вакцинации против инфекционных агентов [13- 15]. Более того, исследования иммуноглобулинов продвинулись вперед, чтобы разработать более безопасные и эффективные методы лечения антителами, что позволяет пересмотреть пассивную иммунизацию для лечения инфекционных заболеваний [16].Наблюдение за иммунологией в этих постоянно разгадывающихся сложностях защиты хозяина снова вернуло нас к В-клеткам в надежде раскрыть ранее недооцененные возможности этих лимфоцитов для улучшения иммунитета против M. tuberculosis .
В- и Т-клетки взаимодействуют для отражения инфекционного заражения
Первыми компонентами защиты организма-хозяина от патогенов с нарушенными анатомическими барьерами являются врожденный иммунитет, который включает мононуклеарные фагоциты, естественные клетки-киллеры и другие врожденные клетки лимфоидного происхождения, нейтрофилы и сывороточные факторы, включая комплемент и естественные антитела [17-19].Встреча с чужеродными микробами через консервативные рецепторы распознавания образов активирует врожденные сторожевые клетки, известные как дендритные клетки, для стимуляции Т-лимфоцитов, которые, в свою очередь, помогают В-клеткам и организуют адаптивные иммунные ответы [20, 21]. Адаптивный иммунитет развился у позвоночных и включает как клеточные, так и гуморальные компоненты, при этом Т-клетки и В-клетки опосредуют эти эффекты соответственно. Т-клетки нацелены и способствуют апоптотическому уничтожению инфицированных патогенами клеток либо напрямую, либо через активацию цитокинами соседних иммунных клеток, в то время как В-клетки вырабатывают антитела, которые нейтрализуют инвазию и нацелены на инфекционные агенты для разрушения [22].Легкие особенно уязвимы для инфекции из-за ограничений анатомических барьеров, обеспечивающих циркуляцию воздуха, что делает эффективный, но непатологический легочный иммунитет жизненно важным для успешной защиты хозяина [23].
Важнейшим из свойств иммунитета является память, которая за счет дифференциации долгоживущих и узкоспециализированных лимфоцитов приводит к определенно эффективным вторичным ответам при последующем воздействии патогенов. Считается, что Эдвард Дженнер первым осознал этот феномен при разработке противооспенной вакцины более 200 лет назад [24].Вакцинация использует иммунологическую память, заряжая иммунную систему ослабленной версией целевого патогена, минуя риск первичной инфекции, сохраняя иммунологические преимущества против будущего контакта с вирулентным патогеном [25]. В качестве эффективного средства предотвращения проникновения патогенов и нейтрализации микробных токсинов антитела имеют особое значение почти для всех вакцин, используемых в настоящее время [26]. Bacillus Calmette-Guérin (БЦЖ) — единственная вакцина, вводимая для профилактики туберкулеза, и, хотя считается, что она защищает от детского туберкулезного менингита, ее эффективность против легочного туберкулеза у взрослых ограничена [27].В одном исследовании эффективность иммунизации БЦЖ против туберкулеза коррелировала со специфическими антителами [28], однако принято считать, что клеточный иммунитет играет преобладающую роль. Поскольку иммунные корреляты защиты против M. tuberculosis остаются не полностью определенными [29], многое еще предстоит понять в отношении способов усиления защиты хозяина от M. tuberculosis . Мы по-прежнему надеемся, что более глубокое понимание как В-, так и Т-клеток будет способствовать улучшению стратегий использования дополнительных функций этих лимфоцитов.
Роль В-клеток в иммунитете против невирусных внутриклеточных патогенов
Защита от внутриклеточных патогенов часто обобщается как исключительно Т-клеточно-опосредованная, при этом В-клетки и антитела играют гораздо более ограниченную роль, чем они воспринимаются во внеклеточной среде. инфекции [30]. Это следует концепции, согласно которой антитела, протекающие через сыворотку, специализируются на нацеливании на внеклеточные микробы, в то время как только Т-клетки, которые используют презентацию антигена как средство поиска внутри клеток, могут специфически нацеливаться на внутриклеточные микробы для уничтожения.Однако существуют исключения из этих правил, поскольку антитела обеспечивают долгоживущую защиту от многих облигатных внутриклеточных вирусов [31-34], предположительно за счет воздействия на внеклеточную фазу вириона, а Т-клетки активируются презентацией антигена, чтобы стимулировать защиту от внеклеточных вирусов. гельминты [35]. Имея прямое отношение к настоящему обзору, появляющиеся экспериментальные данные свидетельствуют о том, что В-клетки играют роль в защите от широкого спектра внутриклеточных бактериальных, грибковых и паразитарных патогенов (см. Ниже).Иммунитет, скорее всего, не развился как два отдельных звена защиты, которые отдельно имеют дело с внутриклеточными и внеклеточными патогенами, скорее, гораздо более изобретательное средство защиты хозяина было разработано для достижения максимально адаптивного, кооперативного и универсального сдерживания инфекции.
Как упоминалось ранее, В- и Т-клеточные ответы дополняют друг друга, отражая естественную инфекцию, и также могут вносить вклад в долгоживущую защиту в результате вакцинации [36, 37]. Например, сотрудничество между клеточным и гуморальным иммунитетом защищает маленьких детей от Haemophilus influenzae типа b, поскольку конъюгация Т-клеточно-независимого углеводного антигена полирибозилрибитолфосфата с Т-клеточно-зависимым белковым иммуногеном стимулирует долгоживущую защиту антител [38, 39 ].Т-хелперы сильно влияют на развитие и специализацию В-клеточных ответов [40], а В-клетки, наоборот, влияют на активацию Т-клеток, действуя как антиген-презентирующие клетки или на них [41]. В рамках этой сложной взаимосвязи между обеими сторонами иммунитета клеточные и гуморальные реакции вместе определяют исход внутриклеточной инфекции.
Вирусы часто полагаются на ограниченные средства клеточного проникновения, а антитела, которые нейтрализуют молекулярные взаимодействия, опосредующие инфекцию, обеспечивают длительный иммунитет против вирусного патогена [42].Внутриклеточные бактерии, такие как микобактерии, и паразиты, такие как малярия, представляют гораздо более сложную картину за счет использования нескольких потенциальных способов проникновения в клетки [43], что увеличивает требования к эпитопам для нейтрализации антител. Учитывая эту сложность, вполне вероятно, что успешные ответы антител против внутриклеточных бактерий имеют тенденцию предотвращать болезнь, а не инфекцию [44], посредством активации путей комплемента и клеточного иммунитета [45]. Необходимы дальнейшие исследования, чтобы понять, как защитные антитела могут быть включены в успешные вакцины против внутриклеточных бактериальных патогенов, таких как M.tuberculosis , которые продолжают вызывать значительную глобальную заболеваемость и смертность.
Ограничение исследований инфекционных заболеваний на мышах с нокаутом одного гена
Небольшой размер, ограниченная стоимость, доступная генетика и лабораторные реактивы, а также иммунная система, которая в значительной степени отражает иммунную систему человека, делают мышей предпочтительным выбором для изучения инфекционных заболеваний. Мыши с целевыми делециями, которые «нокаутируют» определенные гены, являются полезными инструментами для иммунологов и могут использоваться для сбора большого количества информации о роли отдельных иммунных компонентов в ответе хозяина на определенные патогены.Нокаут-мыши были полезны для изучения влияния дефицита В-клеток на прогрессирование заболевания и выживаемость для широкого круга внутриклеточных бактерий и паразитов. Повышенная восприимчивость мышей с дефицитом В-клеток к инфекциям, вызываемым Chlamydia trachomatis , Francisella tularensis , Leishmania major , Plasmodium chabaudi chabaudi , Pneumocystis carinii и , все были зарегистрированы для Salmonella enterica . -52], демонстрируя, что В-лимфоциты опосредуют оптимальный иммунитет против многих внутриклеточных патогенов в дополнение к вирусам.
Исследования инфекции M. tuberculosis у мышей с дефицитом В-клеток были разными, с сообщениями о снижении иммунитета, задержке патологического прогрессирования или отсутствии явных эффектов от генетической абляции В-клеток [10, 11, 53- 55]. Этот широкий диапазон результатов подчеркивает ограничения и различия, присущие мышиным моделям инфекционного заболевания, особенно туберкулеза. Исследования с единичным нокаутом на мышах, инфицированных M. tuberculosis , могут дать результаты, интерпретация которых не является однозначной из-за функционального перекрытия иммунных компонентов [56, 57], может не учитывать различия в ответах потенциальных хозяев между мышами и людьми [58], а также не предоставлять никакой информации о потенциальных эффектах усиления конкретного рассматриваемого иммунного компонента.Следовательно, исследования на мышах с нокаутом могут привести к преждевременным выводам относительно роли определенного компонента иммунитета, если их не интерпретировать тщательно. Кроме того, экспериментальные условия могут иметь заметное влияние на результаты. Например, увеличение инокулята инфекции выявило потребность в В-клетках для оптимального ответа хозяина ТБ у мышей [53, 54]. Подтверждая роль B-клеток в иммунном ответе хозяина на M. tuberculosis , генетическое устранение полимерного рецептора Ig повысило восприимчивость мышей, что указывает на роль секреторного IgA в оптимальном иммунитете против туберкулеза [59].Наконец, защитные эффекты внутривенного иммуноглобулина (ВВИГ) предполагают дальнейшее влияние гуморальных компонентов на защиту хозяина при ТБ [60]. Хотя результаты некоторых исследований на мышах с нокаутом и эксперимента IVIG показывают, что В-клетки и их продукты опосредуют защиту от M. tuberculosis , остается важный вопрос, можно ли усилить ответы В-клеток для повышения иммунитета против M. tuberculosis . с помощью иммунотерапии или вакцинации.
Эктопические агрегаты В-клеток как следствие хронического воспаления и инфекции
Наш новый интерес к В-лимфоцитам возник в результате наблюдений за фолликулоподобными агрегатами В-клеток в легких больных туберкулезом [61, 62].Эти агрегаты В-клеток являются преобладающими очагами клеточной пролиферации в легких человека при ТБ [61] и являются характеристиками гранулематозного прогрессирования ТБ у мышей [11, 62, 63]. Агрегаты В-клеток являются отмеченными патологическими проявлениями многих хронических воспалительных заболеваний, включая рассеянный склероз и ревматоидный артрит [64, 65]. Подобные кластеры В-клеток наблюдались при других инфекциях, таких как вирус гриппа и Helicobacter [66, 67]. Некоторые патогены разработали средства, способствующие размножению и подавлению апоптоза В-клеток [68, 69], которые могут способствовать накоплению этих лимфоцитов во время инфекции.
Каковы эффекты этих эктопических агрегатов В-клеток на иммунитет к M. tuberculosis ? Была выдвинута гипотеза, что эти фолликулоподобные структуры функционируют, чтобы увековечить локальные ответы хозяина, при этом большая часть пролиферативной активности происходит в непосредственной близости от агрегатов В-клеток [61]. Т-клетки могут быть разбросаны по этим кластерам В-клеток [62], создавая эти потенциальные места как презентации антигена, так и созревания В-клеток. Также примечательно, что эти агрегаты В-клеток являются компонентами третичной лимфоидной ткани в легких при туберкулезе [70] и содержат маркеры зародышевых центров [54].Далее предполагая, что В-клетки способствуют локализованной архитектуре иммунного ответа, мы отметили, что у мышей с дефицитом В-клеток обострилась легочная патология и нарушение гранулематозной организации в легких [54]. Еще многое предстоит понять в отношении значения лимфоидного новообразования при ТБ, но было продемонстрировано, что эти индуцируемые лимфоидные структуры могут инициировать защитный иммунитет в легких и реакции памяти против заражения вирусом легочного гриппа в отсутствие вторичных лимфоидных органов [66, 71 ].
На активацию Т-клеток сильно влияют В-клетки.
В-клетки — профессиональные антигенпрезентирующие клетки, оказывающие заметное влияние на ответы Т-клеток. Улавливая антигены через рецепторы клеточной поверхности, инициируется активация В-лимфоцитов. Последующее развитие через клеточные взаимодействия с CD4 + хелперными Т-клетками включает события презентации антигена, которые стимулируют Т-клетки продуцировать цитокины, которые реципрокно регулируют антительный ответ В-клеток [72]. Хотя значение опосредованной В-клетками презентации антигена варьируется в зависимости от антигена и иммунологических условий [73], эти лимфоциты могут быть нацелены на презентацию антигена Т-клеткам с помощью специфических стратегий вакцинации [74, 75].Фактически, один такой вектор вакцины, нацеленный на В-клетки, оказался эффективным в повышении иммунитета, примированного БЦЖ, против M. tuberculosis [76]. В недавно опубликованном примере присутствие В-клеток, но не специфических антител, защищало от реактивации хронической вирусной инфекции, предположительно посредством презентации антигена Т-клеткам [77]. Также сообщалось о независимой от антител функции В-клеток в примировании оптимальных первичных и вторичных иммунных ответов против внутриклеточного бактериального патогена Francisella tularensis [78].Более того, активированные В-клетки, служащие антигенпрезентирующими клетками, используются для усиления противоопухолевого иммунитета [79]. Также было отмечено, что презентация антигена В-клетками может способствовать сохранению аутоиммунитета [80], в то время как покоящиеся В-клетки, как полагают, опосредуют иммунную толерантность [81, 82]. Изучая В-клетки как мишени для усиления иммунитета, а также для подавления аутоиммунитета [83], будущие исследования, вероятно, раскроют творческие средства нацеливания на этот часто упускаемый из виду путь презентации антигена.Наконец, накопленные данные свидетельствуют о том, что В-клетки необходимы для примирования Т-клеточных ответов памяти [52, 84], что помещает взаимосвязь В- и Т-клеток в особенно важный контекст для биологии вакцины.
В-клетки также могут действовать на другие антиген-презентирующие клетки, таким образом влияя на развитие Т-клеточных ответов более косвенным образом. Т-клетки стимулируются к клональной экспансии антиген-презентирующими клетками, которые одновременно экспрессируют костимулирующие молекулы на своей поверхности, одновременно представляя антиген [85].Это иллюстрирует гипотезу о двух сигналах для активации Т-клеток, при которой включение как рецептора антигена, так и CD28 на поверхности Т-клеток необходимо для инициирования адаптивных иммунных ответов, при этом толерантность лимфоцитов является следствием отсутствия второго сигнала [86]. Антигенпрезентирующие клетки активируют поверхностную экспрессию костимулирующих белков, таких как белки семейства B7, во время процесса созревания, который следует за активацией врожденного иммунитета [17]. Эти костимулирующие молекулы затем взаимодействуют с CD28 или другими корецепторами, способствуя активации Т-клеток [87].В-клетки могут модулировать этот процесс созревания за счет выработки антител и цитокинов, которые могут либо усиливать, либо подавлять иммунные ответы [88–93]. Сообщалось, что В-клетки поляризуют ответы Т-клеток за счет продукции цитокинов [94], и обеспечение IL-10 считается средством, с помощью которого В-клетки могут способствовать дифференцировке Th3 у мышей [95]. Кроме того, природные антитела могут модулировать презентацию антигена, связываясь с костимулирующими молекулами B7 и CD40 и изменяя их активность [41, 96, 97].
Многое было обнаружено в отношении регуляции антителами презентации антигена через рецепторы Fcγ [98], и этот иммунологический путь вызвал интерес как потенциальное средство улучшения вакцинации против внутриклеточных патогенов [99]. Рецепторы Fcγ делятся на стимулирующие и ингибирующие типы в зависимости от наличия внутриклеточных ITAM или ITIM-мотивов соответственно [100, 101]. FcγRIIB, единственный ингибирующий рецептор Fcγ, имеет сложное участие в активации Т-клеток, ограничивая созревание дендритных клеток и последующую презентацию антигена, в то время как стимулирующие рецепторы Fcγ, по-видимому, способствуют обоим процессам [89, 90].FcγRIIB также играет важную роль в обеспечении периферической толерантности к ответам Т-клеток в моделях аутоиммунитета мышей [102]. Кроме того, избирательная блокада FcγRIIB приводит к усилению активности Т-клеток в экспериментальных моделях опухолей [89–91]. Стимулирующие рецепторы Fcγ, по-видимому, способствуют ответам Т-клеток, поляризация которых определяется преобладающим воспалительным контекстом и может быть доминирующей либо Th2, либо Th3 [103]. На вовлечение рецептора Fcγ может сильно влиять изотип антитела [104], а методы иммунизации, которые преимущественно нацелены на стимулирующие рецепторы Fcγ, имеют потенциальные эффекты усиления клеточного иммунитета против внутриклеточных патогенов [99, 105].Интересно, что сообщалось, что FcγRIII опосредует иммуносупрессивные эффекты в модели IVIG, иллюстрируя, что ассоциация с мотивом ITAM не может ограничивать функцию провоспалительной активности [106].
Генетическое нарушение функции рецептора Fcγ у мышей повлияло на роль этих рецепторов в микробной инфекции [107]. В частности, специфическое устранение общей стимулирующей Fcγ-цепи нарушает оптимальный иммунитет против множества внутриклеточных патогенов, включая вирус гриппа, видов Leishmania , Plasmodium berghei и Salmonella enterica [49, 108-112].Пассивная иммунизация против Cryptococcus neoformans с помощью IgG1 mAb также зависела от функциональных стимулирующих рецепторов Fcγ [113], что дополнительно задействовало стимулирующие рецепторы Fcγ в защите клетки-хозяина от внутриклеточных патогенов. Мы обнаружили, что нарушение стимулирующей активности рецептора Fcγ увеличивает восприимчивость к M. tuberculosis Erdman, что соответствует нарушению бактериальной локализации и ухудшению иммунопатологии [114]. Напротив, генетическая делеция ингибирующего FcγRIIB улучшает локализацию микобактерий с увеличением продукции IFN-γ и поляризации Th2, обнаруживаемой в легких [114].Предпринимаются попытки изучить, как эти очевидные эффекты рецепторов Fcγ могут быть использованы для повышения иммунитета против M. tuberculosis .
Эти иммуностимулирующие эффекты от специфического нацеливания на рецепторы Fcγ могут обеспечивать механизмы улучшения индуцированного вакциной иммунитета против внутриклеточных патогенов [99, 105]. Например, вирусные векторы могут быть нацелены на клетки, несущие рецептор Fcγ, посредством антителозависимого усиления инфекции [115], обеспечивая средство, с помощью которого можно потенциально манипулировать антигенпрезентирующими клетками для активации и иммунизации.Интересно, что был идентифицирован мерозоитный поверхностный антиген Plasmodium falciparum , который преимущественно индуцирует переключение класса изотипа на IgG2b у мышей [116], подтипа цитофильного иммуноглобулина с преимущественным сродством к стимулирующим рецепторам Fcγ [104]. Дальнейшие исследования должны дополнительно выявить, как рецепторы Fcγ могут быть нацелены и использоваться для повышения иммунитета против внутриклеточных патогенов.
Являются ли антитела защитными против
M. tuberculosis , внутриклеточного патогенаВопреки общепринятому представлению о том, что гуморальный иммунитет незначителен для защиты от туберкулезной палочки, пассивное введение антител показало эффективность против M.tuberculosis с конца XIX века [2]. Однако сывороточная терапия туберкулеза потеряла популярность, когда было обнаружено несоответствие эффективности лечения и препаратов реагентов [117], а появление фармакологических антимикобактериальных средств, таких как стрептомицин, к середине двадцатого века предложило гораздо более надежные вариант [118]. Существуют прекрасные обзоры, касающиеся истории и недавних разработок опосредованного антителами иммунитета к M. tuberculosis [2, 119]. Недавно моноклональные антитела против микобактериального арабиноманнана, гепарин-связывающего гемагглютинина и 16 кДа α-кристаллина продемонстрировали эффективность на моделях туберкулеза у мышей [120–124].Эти антитела опосредуют защиту по-разному, некоторые из них уменьшают микобактериальную нагрузку в тканях, тогда как другие повышают выживаемость животных за счет очевидного уменьшения прогрессирования воспаления [119]. Однако примечателен тот факт, что сообщалось, что вакцина конъюгированной арабиноманнан-белок M. tuberculosis индуцирует более устойчивые ответы антител, чем БЦЖ, но без видимого улучшения выживаемости у мышей [125]. Остается вопрос, является ли мышь оптимальным способом измерения усиленной защиты, учитывая хроническое воспалительное и постоянное инфекционное бремя модели и ограниченные корреляции патологии с человеческим заболеванием.Таким образом, необходимы будущие исследования, чтобы выявить корреляты антител с защитой на моделях людей и животных, если должны быть получены эффективные защитные реакции В-клеток, вызванные иммунизацией, против M. tuberculosis . Наконец, следует отметить, что практически ничего не известно о влиянии врожденных или естественных ответов антител [126-128] на микобактериальную инфекцию. Учитывая, что сложные липиды являются основными составляющими гидрофобной клеточной стенки микобактерий, было бы интересно изучить значение Т-независимых ответов антител, особенно B1 и В-клеток маргинальной зоны, в защите против M.Туберкулез .
Помимо антиген-специфической нейтрализации, антитела также обладают заметными общими эффектами на воспаление, включая активацию комплемента, перекрестное связывание рецептора Fcγ и высвобождение микробных продуктов за счет прямой антимикробной активности [45]. Что касается ответов легочного хозяина, то антитела могут модулировать архитектурные изменения эпителия и сосудов дыхательных путей, как в ответ на инфекцию микоплазмой [129]. В наших собственных исследованиях мы обнаружили, что адоптивный перенос В-клеток устранял обострение воспаления у мышей с дефицитом В-клеток после заражения воздушно-капельным путем M.tuberculosis Эрдман [54]. Предполагая роль опосредованной иммуноглобулином «эндокринной» иммунной регуляции, это снижение патологии происходило в сочетании с обнаруживаемыми уровнями антител в сыворотке, но без локального присутствия В-клеток в легких [54]. Совершенно очевидно, что антитела могут иметь множество защитных эффектов во время инфицирования внутриклеточными патогенами, включая ограничение воспалительной патологии [130] в дополнение к хорошо известной роли в нейтрализации и опсонизации микробов.
Как В-клетки формируют иммунный ответ против
M. tuberculosisКорреляты опосредованной вакциной защиты против M. tuberculosis определены не полностью, но большинство данных свидетельствует о том, что Т-клетки имеют первостепенное значение [7]. Следовательно, цель всех новых противотуберкулезных вакцин, разрабатываемых в настоящее время, заключается в усилении клеточного иммунного ответа против патогена [131]. Как напомнил недавний провал клинических испытаний Т-клеточной вакцины против ВИЧ [132], разумно не ограничивать исследования вакцины слишком узко.Вполне вероятно, что для оптимально успешной иммунизации против M. tuberculosis в будущем потребуются защитные реакции антител [44], и дальнейшие исследования, направленные на раскрытие того, как лучше всего использовать гуморальный иммунитет для обеспечения этой защиты, кажутся оправданными. В этом направлении попытки понять биологию В-клеток и ее связь с туберкулезом показали, что эти лимфоциты, на которые часто не обращают внимания, могут значительно влиять на выработку цитокинов, сдерживание бактерий и иммунопатологическое прогрессирование во время M.tuberculosis инфекция. В качестве одного из возможных механизмов, с помощью которого В-клетки формируют иммунный ответ, мы предполагаем, что иммуноглобулин, действуя на рецепторы Fcγ, влияет на созревание антигенпрезентирующих клеток, вызывая отмеченные фенотипы у мышей с дефицитом генов, зараженных M. tuberculosis (). Однако это лишь один из путей, с помощью которых В-лимфоциты модулируют реакцию хозяина на туберкулезную инфекцию. Предположительно, В-клетки могут формировать противотуберкулезный иммунитет различными способами, включая прямое воздействие антител на патоген, презентацию антигена, продукцию цитокинов, а также влияние на механизмы внутриклеточного уничтожения лейкоцитов.
Схема того, как B-клетки формируют иммунный ответ против M. tuberculosisB-клетки модулируют ответ мыши-хозяина против M. tuberculosis различными способами. При острой инфекции избирательное включение стимулирующих рецепторов Fcγ комплексами антител усиливает ответ Th2 и способствует сдерживанию микобактерий с минимальным воспалением. Напротив, участие ингибирующего FcγRIIB увеличивает продукцию IL-10 и ставит под угрозу иммунитет против M.Туберкулез . Интересно, что иммуносупрессивный фенотип в отсутствие В-клеток изначально нарушает оптимальное сдерживание острого заражения M. tuberculosis , но задерживает прогрессирование воспаления при хроническом ТБ.
Как указывалось ранее, активация Т-клеток достигается посредством презентации антигена специализированными клетками, которые должны пройти процесс созревания, чтобы выполнять эту функцию [17]. Ранее мы описали, как В-клетки могут оказывать значительное влияние на созревание антигенпрезентирующих клеток за счет взаимодействия с рецепторами Fcγ антителами.Используя мышей с дефицитом ингибирующей функции рецептора FcγRIIB, мы обнаружили, что отсутствие FcγRIIB усиливает сдерживание M. tuberculosis Erdman у мышей [114]. Продукция легочного IFN-γ и частота клеток Th2 соответственно увеличивались у FcγRIIB-дефицитных мышей. Напротив, генетическое устранение общей γ-цепи, общей для стимулирующих рецепторов Fcγ, нарушает локализацию M. tuberculosis и ухудшает общую выживаемость (110). Таким образом, В-клетки могут значительно влиять на иммунитет хозяина и исход заболевания за счет вовлечения рецепторов Fcγ во время ТБ, влияя как на активацию Th2, так и на сдерживание микобактерий.
Исследования на мышах инфекции M. tuberculosis также показывают, что В-клетки оказывают значительное влияние на продукцию IL-10 в легких. ИЛ-10 продуцируется широким спектром лейкоцитов, включая В-клетки, дендритные клетки, макрофаги и Т-клетки, выполняя в основном противовоспалительные функции в ответах хозяина [133]. Повышенная продукция IL-10 в легких во время инфекции M. tuberculosis была обнаружена в двух различных моделях дефицита В-клеток на мышах [54, 134].Мы также наблюдали увеличение IL-10 в легких мышей с дефицитом стимулирующей Fcγ-цепи, инфицированных M. tuberculosis Erdman [114].
Клеточный источник этого увеличения IL-10 неясен, поскольку методы окрашивания внутриклеточных цитокинов для обнаружения IL-10 не являются оптимально чувствительными для обнаружения значительных количеств этих клеток у мышей. Недавнее развитие мышей-репортеров IL-10 может облегчить такие исследования в будущем [135, 136]. Продукция IL-10 характерна для не полностью активированных дендритных клеток, которые не претерпели полного созревания [137].Таким образом, влияя на активацию клеток через взаимодействие иммунных комплексов с рецепторами Fcγ, В-клетки могут влиять на продукцию IL-10 этими антигенпрезентирующими клетками. Также сообщалось, что В-клетки могут активировать или ингибировать регуляторные Т-клетки, важный клеточный источник ИЛ-10 [138, 139]. Наконец, повышение уровня IL-10 может быть реакцией на обострение иммунопатологии, индуцированной во время инфицирования M. tuberculosis B-клеток — / — и Fcγ-цепи — / — мышей [54], поскольку производство противовоспалительных цитокинов , как и IL-10, может быть компенсирующей попыткой ограничить чрезмерное воспаление.Необходима дополнительная работа для оценки взаимосвязи В-клеток и цитокинов во время туберкулеза, особенно с учетом того, что сами В-клетки могут быть источником жизненно важного антимикобактериального цитокина IFN-γ [94].
Как также описано ранее в этой статье, В-клетки являются важными компонентами ТБ легочного гранулематозного воспаления с большими агрегатами этих лимфоцитов как характерными гистологическими характеристиками инфекции [61-63]. Отсутствие В-клеток при одновременном нарушении оптимального иммунитета приводит к обострению легочной патологии при острой болезни M.tuberculosis [54], парадоксальным образом приводит к задержке прогрессирования воспаления во время хронической инфекции у мышей [55]. Одно из возможных объяснений этого парадокса состоит в том, что функция В-клеток во время туберкулезной инфекции зависит от фазы инфекции: во время острой инфекции В-клетки необходимы для оптимального гранулематозного ответа и эффективного иммунитета против легочного заражения M. tubeculosis и в их отсутствие требуется нарушение регуляции образования гранулем и усиление легочного воспаления для сдерживания инфекции.Напротив, во время хронической фазы инфекции, когда содержатся устойчивые бациллы, иммунологически активные кластеры В-клеток [54, 61, 62], вероятно, способствуют сохранению местного иммунитета хозяина против M. tuberculosis и могут помочь в предотвращении реактивации болезни. Это продолжение воспаления может происходить частично за счет В-клеток, действующих как антигенпрезентирующие клетки. Действительно, было обнаружено, что Т-клетки связаны с узелками В-клеток в туберкулезных тканях легких как человека, так и мыши ([62], P.Maglione и J. Chan, неопубликовано). В этом отношении захват антигена может происходить особым образом через BCR, хотя было показано, что B-клетки могут загружать MHC пептидом не в соответствии со специфичностью их рецептора [140]. Такие «неспецифические» антигены могут быть результатом поглощения Toll-подобным рецептором [141]. Таким образом, воспалительный парадокс мышей с дефицитом В-клеток, по-видимому, отражает роль В-клеток, переходящую от оптимизации защиты хозяина во время острого заражения к сохранению хронической воспалительной реакции во время персистирующей инфекции.
Заболеваемость и смертность от туберкулеза является результатом аберрантной и повреждающей реакции хозяина [142, 143]: она является чрезмерной по патологическим последствиям, но неэффективна в сдерживании патогена. Этот очевидный парадокс можно рассматривать как результат неэффективного иммунного сдерживания туберкулезной палочки, ведущего к чрезмерному компенсаторному рекрутированию лейкоцитов в легких. Лучше всего это проявляется у пациентов с реактивацией ТБ. Вместо того, чтобы быть лишенными иммунной активности, которая могла бы объяснить повторное появление спящего патогена, легкие пациентов с реактивационными заболеваниями содержат области интенсивного легочного инфильтрата [143].До лечения в активном легочном инфильтрате ТБ преобладают нейтрофилы [61], врожденная иммунная клетка, которая способствует ранней защите, но способствует воспалительному повреждению при различных острых легочных заболеваниях [144]. Как и у людей, восприимчивость к туберкулезу связана с повышенным притоком нейтрофилов в легкие у мышей [145]. Напротив, случайные находки комплексной патологии Гона у людей подчеркивают, что для успешного сдерживания M. tuberculosis не требуется значительная иммунопатология в качестве побочного продукта.Существующие данные убедительно свидетельствуют о том, что В-клетки играют роль в модулировании воспалительной реакции во время туберкулезной инфекции [54,55]
Заключительные замечания
Даже если сдерживание бактерий хозяина проходит без симптомов болезни, инфекция M. tuberculosis сохраняется в латентной форме. состояние, которое может реактивироваться в более позднем возрасте, часто в результате иммунодефицита. Учитывая это наблюдение, что инфекция M. tuberculosis сохраняется даже у тех, у кого есть успешный профилактический иммунитет, ясно, что вакцина, предотвращающая туберкулез, должна вызывать иммунный ответ, превосходящий иммунный ответ, вызванный естественной инфекцией [44].Рациональный дизайн такой вакцины потребует обширного изучения базовой иммунологии, лежащей в основе защитного иммунитета, и, вероятно, выиграет от понимания того, как иммунологические пути, которые могут быть умеренно защитными во время естественной инфекции, могут быть оптимизированы для достижения нейтрализации посредством вакцинации.
Поскольку этот обзор направлен на повышение интереса к взаимосвязи биологии В-клеток и туберкулеза, важно выделить ключевые вопросы, на которые необходимо ответить. Например, какие типы В-клеток памяти развиваются, если вообще развиваются, в результате естественных M.туберкулез инфекция? Как эти В-клетки памяти сравниваются с теми, которые были вызваны вакцинацией БЦЖ? Развиваются ли Т-независимые В-клетки памяти против небелковых антигенов M. tuberculosis , и являются ли они лучшими мишенями для иммунизации, чем белковые антигены? Кроме того, можно ли идентифицировать антитела, которые предотвращают микобактериальную инфекцию у людей, и можно ли включить эти мишени нейтрализации в вакцину? И наоборот, существуют ли специфические антитела, которые отрицательно влияют на противотуберкулезный иммунитет? Обнаружены ли эктопические зародышевые центры в легких при туберкулезе важными участками развития В-клеток памяти или долгоживущих плазматических клеток? Какова патология скрытой инфекции и присутствуют ли зародышевые центры на протяжении всего течения хронического туберкулеза? Можно ли стимулировать В-клетки в качестве альтернативы стерильному уничтожению для поддержания хронического сдерживания и предотвращения реактивации ТБ? Или, наоборот, можно ли использовать лечение истощения B-клеток для уменьшения воспалительной патологии у пациентов с хроническим туберкулезом? Какие антитело-независимые функции В-клеток участвуют в формировании иммунного ответа хозяина на M.туберкулез ?
Многие вопросы о роли В-клеток остаются без ответа, но мы надеемся, что дальнейшие исследования этих лимфоцитов покажут больше о том, как их можно использовать для оптимизации иммунитета против M. tuberculosis . Учитывая глобальное значение туберкулеза, крайне важно, чтобы мы исследовали все возможности, которые могут помочь в разработке более эффективных методов лечения и разработке эффективных вакцин. Несмотря на многолетние представления о бесполезности В-клеток по отношению к инфекции M. tuberculosis , недавние данные показывают, что нельзя упускать из виду это мощное звено иммунитета.
Выражение признательности
Эта работа была поддержана грантами Национального института здравоохранения R01 HL071241, R01 AI50732, P01 AI063537 и Медицинским колледжем Альберта Эйнштейна / Медицинским центром Монтефиоре по исследованию СПИДа, грантами P30 AI051519 и T32 AI007506, а также посевным грантом. от Фонда Американской медицинской ассоциации (в PJM).
Сокращения
- TB
- tuberculosis
- IVIG
- внутривенный иммуноглобулин
- Ig
- иммуноглобулин
Ссылки
2.Глатман-Фридман А., Касадевалл А. Еще раз о сывороточной терапии туберкулеза: переоценка роли опосредованного антителами иммунитета против Mycobacterium tuberculosis. Clin Microbiol Rev.1998; 11: 514-532. [Бесплатная статья PMC] [PubMed] [Google Scholar] 3. Исеман MD. Туберкулезная терапия: прошлое, настоящее и будущее. Eur Respir J Suppl. 2002; 36: 87с – 94с. [PubMed] [Google Scholar] 4. Germain RN. Врожденно интересное десятилетие исследований в области иммунологии. Nat Med. 2004. 10: 1307–1320. [PubMed] [Google Scholar] 5. Кальметт А.Пассивный иммунитет: попытки серотерапии против туберкулеза. The Williams & Wilkins Co .; Балтимор, Мэриленд: 1923. [Google Scholar] 6. Сильверштейн AM. Дарвинизм и иммунология: от Мечникова до Бернета. Nat Immunol. 2003; 4: 3–6. [PubMed] [Google Scholar] 7. Флинн Дж. Л., Чан Дж. Иммунология туберкулеза. Анну Рев Иммунол. 2001; 19: 93–129. [PubMed] [Google Scholar] 8. Казанова Дж. Л., Абель Л. Генетическое вскрытие иммунитета к микобактериям: модель человека. Анну Рев Иммунол. 2002. 20: 581–620. [PubMed] [Google Scholar] 9.Doffinger R, Patel S, Kumararatne DS. Иммунодефициты человека, предрасполагающие к внутриклеточным бактериальным инфекциям. Curr Opin Rheumatol. 2005. 17: 440–446. [PubMed] [Google Scholar] 10. Джонсон С.М., Купер А.М., Фрэнк А.А., Бонорино С.Б., Высоки Л.Дж., Орм И.М. Инфекции, вызываемые аэрогенными микобактериями туберкулеза у мышей с дефицитом В-клеток. Tuber Lung Dis. 1997. 78: 257–261. [PubMed] [Google Scholar] 11. Тернер Дж., Фрэнк А.А., Брукс Дж. В., Гонсалес-Хуарреро М., Орм И.М. Развитие хронического туберкулеза у мышей не требует участия В-лимфоцитов или интерлейкина-4.Exp Gerontol. 2001; 36: 537–545. [PubMed] [Google Scholar] 12. Интлекофер А.М., Джон Уэрри Э., Райнер С.Л. Не очень большие ожидания: переоценка сущности Т-клеточной памяти. Immunol Rev.2006; 211: 203–213. [PubMed] [Google Scholar] 14. Пулендран Б., Ахмед Р. Перевод врожденного иммунитета в иммунологическую память: значение для разработки вакцины. Клетка. 2006; 124: 849–863. [PubMed] [Google Scholar] 15. Кауфманн С.Х., Бауманн С., Нассер Эддин А. Использование иммунологии и молекулярной генетики для рационального дизайна вакцины против туберкулеза.Int J Tuberc Lung Dis. 2006; 10: 1068–1079. [PubMed] [Google Scholar] 16. Casadevall A, Dadachova E, Pirofski LA. Пассивная терапия антителами при инфекционных заболеваниях. Nat Rev Microbiol. 2004. 2: 695–703. [PubMed] [Google Scholar] 17. Меджитов Р., Джейнвей С. Младший Врожденный иммунитет. N Engl J Med. 2000; 343: 338–344. [PubMed] [Google Scholar] 18. Tosi MF. Врожденный иммунный ответ на инфекцию. J Allergy Clin Immunol. 2005; 116: 241–249. викторина 250. [PubMed] [Google Scholar] 19. Браун KL, Cosseau C, Гарди JL, Hancock RE.Сложности нацеливания на врожденный иммунитет для лечения инфекции. Trends Immunol. 2007. 28: 260–266. [PubMed] [Google Scholar] 20. Меджитов Р., Престон-Херлбурт П., Джейнвей К.А., мл. Человеческий гомолог белка Toll дрозофилы сигнализирует об активации адаптивного иммунитета. Природа. 1997. 388: 394–397. [PubMed] [Google Scholar] 21. Меджитов Р., Джейнвей С., мл. Семейство рецепторов Толля и распознавание микробов. Trends Microbiol. 2000. 8: 452–456. [PubMed] [Google Scholar] 22. Мур BB, Мур Т.А., Тэйвс ГБ. Роль Т- и В-лимфоцитов в легочной защите хозяина.Eur Respir J. 2001; 18: 846–856. [PubMed] [Google Scholar] 23. Чжан П., Саммер В.Р., Бэгби Г.Дж., Нельсон С. Врожденный иммунитет и легочная защита хозяина. Immunol Rev.2000; 173: 39–51. [PubMed] [Google Scholar] 24. Волленберг А., Энглер Р. Оспа, вакцинация и побочные реакции на противооспенную вакцину. Curr Opin Allergy Clin Immunol. 2004; 4: 271–275. [PubMed] [Google Scholar] 25. Джейнвей Калифорния младший, Трэверс П., Уолпорт М., Шломчик М.Дж. Иммунобиология: иммунная система в здоровье и болезнях. Издательство Garland; Нью-Йорк, Нью-Йорк: 2001.[Google Scholar] 26. Gourley TS, Wherry EJ, Masopust D, Ahmed R. Создание и поддержание иммунологической памяти. Semin Immunol. 2004. 16: 323–333. [PubMed] [Google Scholar] 27. Сандер С., МакШейн Х. Серия трансляционных мини-обзоров вакцин: разработка и оценка улучшенных вакцин против туберкулеза. Clin Exp Immunol. 2007. 147: 401–411. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Костелло А.М., Кумар А., Нараян В., Акбар М.С., Ахмед С., Абу-Зейд С., Рук Г.А., Стэнфорд Дж., Морено С. Ограничивают ли антитела к микобактериальным антигенам, включая липоарабиноманнан, распространение туберкулеза у детей? Trans R Soc Trop Med Hyg.1992. 86: 686–692. [PubMed] [Google Scholar] 29. Hanekom WA. Иммунный ответ новорожденных на вакцинацию БЦЖ. Ann N Y Acad Sci. 2005; 1062: 69–78. [PubMed] [Google Scholar] 30. Седер Р.А., Хилл А.В. Вакцины против внутриклеточных инфекций, требующие клеточного иммунитета. Природа. 2000; 406: 793–798. [PubMed] [Google Scholar] 31. Даудл В. Р., Дауни Дж. С., Лейвер В. Г.. Подавление высвобождения вируса антителами к поверхностным антигенам вирусов гриппа. J Virol. 1974; 13: 269–275. [Бесплатная статья PMC] [PubMed] [Google Scholar] 32.Fujinami RS, Oldstone MB. Противовирусные антитела, реагирующие на плазматическую мембрану, изменяют экспрессию вируса кори внутри клетки. Природа. 1979; 279: 529–530. [PubMed] [Google Scholar] 33. Левин Б., Хардвик Дж. М., Трапп Б. Д., Кроуфорд ТО, Боллинджер Р. К., Гриффин Д. Опосредованное антителами очищение нейронов от альфавирусной инфекции. Наука. 1991; 254: 856–860. [PubMed] [Google Scholar] 34. Родригес Дж. Э., Монингер Т., Гроуз С. Вход и выход вируса ветряной оспы блокируются одним и тем же моноклональным антителом против gH. Вирусология.1993; 196: 840–844. [PubMed] [Google Scholar] 35. Рамалингам Т.Р., Рейман Р.М., Винн Т.А. Использование моделей гельминтов и аллергии для понимания биологии цитокинов Th3. Curr Opin Allergy Clin Immunol. 2005; 5: 392–398. [PubMed] [Google Scholar] 36. Доэрти П.С., Тернер С.Дж., Уэбби Р.Г., Томас П.Г. Грипп и вызов иммунологии. Nat Immunol. 2006. 7: 449–455. [PubMed] [Google Scholar] 37. Аманна И.Дж., Слифка М.К., Кротти С. Иммунитет и иммунологическая память после вакцинации против оспы. Immunol Rev.2006; 211: 320–337.[PubMed] [Google Scholar] 38. Шнеерсон Р., Баррера О., Саттон А., Роббинс Дж. Б. Получение, характеристика и иммуногенность конъюгатов полисахарид-белок Haemophilus influenzae типа b. J Exp Med. 1980; 152: 361–376. [Бесплатная статья PMC] [PubMed] [Google Scholar] 39. Деккер MD, Эдвардс KM. Вакцины против Haemophilus influenzae типа b: история болезни, выбор и сравнение. Pediatr Infect Dis J. 1998; 17: S113–116. [PubMed] [Google Scholar] 40. Винуэса К.Г., Танге С.Г., Мозер Б., Маккей С.Р. Фолликулярные В-хелперные Т-клетки в ответах антител и аутоиммунитете.Nat Rev Immunol. 2005; 5: 853–865. [PubMed] [Google Scholar] 41. Bayry J, Lacroix-Desmazes S, Kazatchkine MD, Hermine O, Tough DF, Kaveri SV. Модуляция созревания и функции дендритных клеток с помощью В-лимфоцитов. J Immunol. 2005; 175: 15–20. [PubMed] [Google Scholar] 42. Бертон ДР. Антитела, вирусы и вакцины. Nat Rev Immunol. 2002; 2: 706–713. [PubMed] [Google Scholar] 43. Писарро-Серда Дж., Коссарт П. Бактериальная адгезия и проникновение в клетки-хозяева. Клетка. 2006; 124: 715–727. [PubMed] [Google Scholar] 44. Кауфманн Ш.Вклад иммунологии в рациональный дизайн новых антибактериальных вакцин. Nat Rev Microbiol. 2007; 5: 491–504. [PubMed] [Google Scholar] 45. Casadevall A, Pirofski LA. Переоценка гуморального иммунитета на основе механизмов опосредованной антителами защиты от внутриклеточных патогенов. Adv Immunol. 2006; 91: 1–44. [PubMed] [Google Scholar] 46. Су Х, Фейлзер К., Колдуэлл HD, Моррисон РП. Инфекция половых путей Chlamydia trachomatis у мышей с нокаутом гена с дефицитом антител. Infect Immun. 1997; 65: 1993–1999.[Бесплатная статья PMC] [PubMed] [Google Scholar] 47. Ян X, Brunham RC. Мыши с дефицитом В-клеток с нокаутом гена демонстрируют, что В-клетки играют важную роль в инициации Т-клеточного ответа на инфекцию легких Chlamydia trachomatis (мышиный пневмонит). J Immunol. 1998. 161: 1439–1446. [PubMed] [Google Scholar] 48. Калкин С.Дж., Райнхарт-Джонс Т., Элкинс К.Л. Новая роль В-клеток в раннем защитном иммунитете к внутриклеточному патогену, штамму LVS Francisella tularensis. J Immunol. 1997; 158: 3277–3284.[PubMed] [Google Scholar] 49. Woelbing F, Kostka SL, Moelle K, Belkaid Y, Sunderkoetter C, Verbeek S, Waisman A, Nigg AP, Knop J, Udey MC, von Stebut E.Поглощение Leishmania major дендритными клетками опосредуется рецепторами Fcgamma и способствует приобретению защитный иммунитет. J Exp Med. 2006; 203: 177–188. [Бесплатная статья PMC] [PubMed] [Google Scholar] 50. Langhorne J, Cross C, Seixas E, Li C, von der Weid T. Роль B-клеток в развитии функции T-хелперов при малярийной инфекции у мышей.Proc Natl Acad Sci U S. A. 1998; 95: 1730–1734. [Бесплатная статья PMC] [PubMed] [Google Scholar] 51. Mastroeni P, Simmons C, Fowler R, Hormaeche CE, Dougan G. Мыши Igh-6 (- / -) (с дефицитом B-клеток) не могут сформировать устойчивую приобретенную устойчивость к пероральному заражению вирулентным сероварным тифимурием Salmonella enterica и демонстрируют нарушение Th2 Т-клеточные ответы на антигены сальмонелл. Infect Immun. 2000. 68: 46–53. [Бесплатная статья PMC] [PubMed] [Google Scholar] 52. Lund FE, Hollifield M, Schuer K, Lines JL, Randall TD, Garvy BA.В-клетки необходимы для генерации защитных эффекторных клеток и клеток памяти CD4 в ответ на инфекцию легких, вызванную пневмоцистами. J Immunol. 2006. 176: 6147–6154. [PubMed] [Google Scholar] 53. Vordermeier HM, Venkataprasad N, Harris DP, Ivanyi J. Увеличение туберкулезной инфекции в органах мышей с дефицитом B-клеток. Clin Exp Immunol. 1996. 106: 312–316. [Бесплатная статья PMC] [PubMed] [Google Scholar] 54. Маглионе П.Дж., Сюй Дж., Чан Дж. В-клетки замедляют прогрессирование воспаления и усиливают сдерживание бактерий при легочном заражении Mycobacterium tuberculosis.J Immunol. 2007; 178: 7222–7234. [PubMed] [Google Scholar] 55. Bosio CM, Gardner D, Elkins KL. Заражение мышей с дефицитом В-клеток CDC 1551, клиническим изолятом Mycobacterium tuberculosis: задержка распространения и развития патологии легких. J Immunol. 2000. 164: 6417–6425. [PubMed] [Google Scholar] 56. Скотт HM, Флинн JL. Mycobacterium tuberculosis у мышей с дефицитом хемокинового рецептора 2: влияние дозы на прогрессирование заболевания. Infect Immun. 2002; 70: 5946–5954. [Бесплатная статья PMC] [PubMed] [Google Scholar] 57.Bafica A, Scanga CA, Feng CG, Leifer C, Cheever A, Sher A. TLR9 регулирует ответы Th2 и взаимодействует с TLR2, обеспечивая оптимальную устойчивость к Mycobacterium tuberculosis. J Exp Med. 2005; 202: 1715–1724. [Бесплатная статья PMC] [PubMed] [Google Scholar] 58. Флинн JL. Уроки экспериментальных инфекций Mycobacterium tuberculosis. Микробы заражают. 2006; 8: 1179–1188. [PubMed] [Google Scholar] 59. Tjarnlund A, Rodriguez A, Cardona PJ, Guirado E, Ivanyi J, Singh M, Troye-Blomberg M, Fernandez C. Мыши с нокаутом полимерного IgR более восприимчивы к микобактериальным инфекциям дыхательных путей, чем мыши дикого типа.Int Immunol. 2006. 18: 807–816. [PubMed] [Google Scholar] 60. Рой Е., Ставропулос Е., Бреннан Дж., Коуд С., Григорьева Е., Уокер Б., Дагг Б., Таскон Р. Э., Лоури Д. Б., Колстон М. Дж., Джоллес С. Терапевтическая эффективность высоких доз внутривенного иммуноглобулина при инфекции Mycobacterium tuberculosis у мышей. Infect Immun. 2005. 73: 6101–6109. [Бесплатная статья PMC] [PubMed] [Google Scholar] 61. Ульрихс Т., Космиади Г.А., Трусов В., Йорг С., Прадл Л., Титухина М., Мищенко В., Гушина Н., Кауфманн Ш. Туберкулезные гранулемы человека индуцируют периферические лимфоидные фолликулоподобные структуры, которые организуют локальную защиту хозяина в легких.J Pathol. 2004. 204: 217–228. [PubMed] [Google Scholar] 62. Tsai MC, Chakravarty S, Zhu G, Xu J, Tanaka K, Koch C, Tufariello J, Flynn J, Chan J. Характеристика туберкулезной гранулемы в легких мыши и человека: клеточный состав и относительное напряжение кислорода в тканях. Cell Microbiol. 2006; 8: 218–232. [PubMed] [Google Scholar] 63. Гонсалес-Хуарреро М., Тернер О.К., Тернер Дж., Мариетта П., Брукс СП, Орм И.М. Временное и пространственное расположение лимфоцитов в гранулемах легких, вызванных аэрозольной инфекцией Mycobacterium tuberculosis.Infect Immun. 2001; 69: 1722–1728. [Бесплатная статья PMC] [PubMed] [Google Scholar] 64. Серафини Б., Розикарелли Б., Маглиоцци Р., Стиглиано Э., Алоизи Ф. Обнаружение эктопических В-клеточных фолликулов с зародышевыми центрами в мозговых оболочках пациентов с вторичным прогрессирующим рассеянным склерозом. Brain Pathol. 2004. 14: 164–174. [Бесплатная статья PMC] [PubMed] [Google Scholar] 65. Manzo A, Paoletti S, Carulli M, Blades MC, Barone F, Yanni G, Fitzgerald O, Bresnihan B, Caporali R, Montecucco C, Uguccioni M, Pitzalis C.Систематический микроанатомический анализ продукции CXCL13 и CCL21 in situ и прогрессивной лимфоидной организации при ревматоидном синовите.Eur J Immunol. 2005; 35: 1347–1359. [PubMed] [Google Scholar] 66. Мойрон-Кирос Дж. Э., Ранжел-Морено Дж., Куссер К., Хартсон Л., Спраг Ф., Гудрич С., Вудленд Д. Л., Лунд Ф. Е., Рэндалл Т. Д.. Роль индуцибельной лимфоидной ткани, ассоциированной с бронхом (iBALT), в респираторном иммунитете. Nat Med. 2004; 10: 927–934. [PubMed] [Google Scholar] 67. Шомер NH, Fox JG, Juedes AE, Ruddle NH. Хеликобактер-индуцированные хронические активные лимфоидные агрегаты имеют характеристики третичной лимфоидной ткани. Infect Immun. 2003. 71: 3572–3577. [Бесплатная статья PMC] [PubMed] [Google Scholar] 68.Бюссьер Ф. И., Чатурведи Р., Асим М., Хук К. Л., Ченг Й., Гейнор Дж., Шольц А., Хан В. Н., Уилсон К. Т.. Низкая множественность заражения Helicobacter pylori подавляет апоптоз В-лимфоцитов. Cancer Res. 2006; 66: 6834–6842. [PubMed] [Google Scholar] 69. Spera JM, Ugalde JE, Mucci J, Comerci DJ, Ugalde RA. Митоген B-лимфоцитов — это фактор вирулентности Brucella abortus, необходимый для стойкой инфекции. Proc Natl Acad Sci U S. A. 2006; 103: 16514–16519. [Бесплатная статья PMC] [PubMed] [Google Scholar] 70. Канерт А, Хопкен У. Э., Штейн М, Бандерманн С, Липп М, Кауфманн Ш.Mycobacterium tuberculosis вызывает образование лимфоидной структуры в легких мышей. J Infect Dis. 2007; 195: 46–54. [PubMed] [Google Scholar] 71. Мойрон-Квироз Дж. Э., Ранжель-Морено Дж., Хартсон Л., Куссер К., Тайге М. П., Клоновски К. Д., Лефрансуа Л., Каули Л. С., Хармсен А. Г., Лунд Ф. Е., Рэндалл Т. Д.. Сохранение и отзывчивость иммунологической памяти при отсутствии вторичных лимфоидных органов. Иммунитет. 2006. 25: 643–654. [PubMed] [Google Scholar] 72. Vascotto F, Le Roux D, Lankar D, Faure-Andre G, Vargas P, Guermonprez P, Lennon-Dumenil AM.Презентация антигена В-лимфоцитами: как передача сигналов рецептора направляет движение через мембрану. Curr Opin Immunol. 2007; 19: 93–98. [PubMed] [Google Scholar] 73. Родригес-Пинто Д. В-клетки как антигенпрезентирующие клетки. Cell Immunol. 2005. 238: 67–75. [PubMed] [Google Scholar] 74. Hon H, Oran A, Brocker T, Jacob J. B лимфоциты участвуют в перекрестной презентации антигена после вакцинации генным пистолетом. J Immunol. 2005; 174: 5233–5242. [PubMed] [Google Scholar] 75. Хелгеби А., Робсон Н. С., Доначи А. М., Бекок-Шарп Х., Ловгрен К., Шон К., Моват А., Лик Нью-Йорк.Комбинированный адъювантный вектор CTA1-DD / ISCOM способствует примированию слизистой оболочки и системному иммунитету к включенным антигенам путем специфического нацеливания на В-клетки. J Immunol. 2006. 176: 3697–3706. [PubMed] [Google Scholar] 76. Andersen CS, Dietrich J, Agger EM, Lycke NY, Lovgren K, Andersen P. Комбинированный вектор CTA1-DD / ISCOMs представляет собой эффективный интраназальный адъювант для усиления предшествующего иммунитета Mycobacterium bovis BCG к Mycobacterium tuberculosis. Infect Immun. 2007. 75: 408–416. [Бесплатная статья PMC] [PubMed] [Google Scholar] 77.McClellan KB, Gangappa S, Speck SH, Virgin HWt. Антитело-независимый контроль латентности гамма-герпесвируса посредством индукции В-клетками противовирусных Т-клеточных ответов. PLoS Pathog. 2006; 2: e58. [Бесплатная статья PMC] [PubMed] [Google Scholar] 78. Элкинс К.Л., Босио К.М., Райнхарт-Джонс ТР. Важность В-клеток, но не специфических антител, в первичном и вторичном защитном иммунитете к внутриклеточному штамму живой вакцины Francisella tularensis. Infect Immun. 1999. 67: 6002–6007. [Бесплатная статья PMC] [PubMed] [Google Scholar] 79.Schultze JL, Grabbe S, von Bergwelt-Baildon MS. DC и CD40-активированные B-клетки: текущие и будущие направления иммунотерапии клеточного рака. Trends Immunol. 2004. 25: 659–664. [PubMed] [Google Scholar] 80. Чан ОТ, Ханнум Л.Г., Хаберман А.М., Мадайо М.П., Шломчик М.Дж. Новая мышь с В-клетками, но без сывороточных антител, обнаруживает антитело-независимую роль В-клеток в мышиной волчанке. J Exp Med. 1999; 189: 1639–1648. [Бесплатная статья PMC] [PubMed] [Google Scholar] 81. Цитура Д.К., Йунг В.П., ДеКрюфф Р.Х., Умецу Д.Т.Критическая роль В-клеток в развитии толерантности Т-клеток к аэроаллергенам. Int Immunol. 2002. 14: 659–667. [PubMed] [Google Scholar] 82. Чен X, Jensen PE. Передний край: первичные В-лимфоциты преимущественно увеличивают аллогенные Т-клетки FoxP3 + CD4. J Immunol. 2007; 179: 2046–2050. [PubMed] [Google Scholar] 83. Hackett CJ, Rotrosen D, Auchincloss H, Fauci AS. Иммунологические исследования: проблемы и возможности в условиях бюджетных ограничений. Nat Immunol. 2007. 8: 114–117. [PubMed] [Google Scholar] 84.Шен Х, Уитмир Дж. К., Фан Х, Шедлок Д. Д., Кеч С. М., Ахмед Р. Особая роль В-клеток в генерации Т-клеточной памяти CD8 рекомбинантными Listeria monocytogenes. J Immunol. 2003; 170: 1443–1451. [PubMed] [Google Scholar] 85. Liu Y, Janeway CA., Jr. Клетки, которые обладают как специфической лигандной, так и костимулирующей активностью, являются наиболее эффективными индукторами клональной экспансии нормальных CD4 Т-клеток. Proc Natl Acad Sci U S. A. 1992; 89: 3845–3849. [Бесплатная статья PMC] [PubMed] [Google Scholar] 86. Джейнвей Калифорния, младший Путешествие по моей жизни на иммунологическую тему.Анну Рев Иммунол. 2002; 20: 1–28. [PubMed] [Google Scholar] 87. Шарп А.Х., Фриман Г.Дж. Суперсемейство B7-CD28. Nat Rev Immunol. 2002; 2: 116–126. [PubMed] [Google Scholar] 89. Калергис А.М., Равеч СП. Индуцирование опухолевого иммунитета за счет избирательного взаимодействия активирующих рецепторов Fcgamma на дендритных клетках. J Exp Med. 2002; 195: 1653–1659. [Бесплатная статья PMC] [PubMed] [Google Scholar] 90. Рафик К., Бергтольд А., Клайнс Р. Представление антигена, опосредованное иммунным комплексом, индуцирует опухолевый иммунитет. J Clin Invest. 2002; 110: 71–79.[Бесплатная статья PMC] [PubMed] [Google Scholar] 91. Dhodapkar KM, Kaufman JL, Ehlers M, Banerjee DK, Bonvini E, Koenig S, Steinman RM, Ravetch JV, Dhodapkar MV. Селективная блокада ингибирующего рецептора Fcgamma делает возможным созревание дендритных клеток человека с выработкой IL-12p70 и иммунитет к опухолевым клеткам, покрытым антителами. Proc Natl Acad Sci U S. A. 2005; 102: 2910–2915. [Бесплатная статья PMC] [PubMed] [Google Scholar] 92. Мидзогути А., Огава А., Такедацу Х., Сугимото К., Шимомура Ю., Ширане К., Нагахама К., Нагайши Т., Мидзогути Е., Блумберг Р.С., Бхан А.К.Зависимость образования гранулемы кишечника от уникальных миелоидных DC-подобных клеток. J Clin Invest. 2007. 117: 605–615. [Бесплатная статья PMC] [PubMed] [Google Scholar] 93. Сугимото К., Огава А., Шимомура Ю., Нагахама К., Мидзогути А., Бхан А. К.. Индуцируемые В-клетки, продуцирующие IL-12, регулируют Th3-опосредованное воспаление кишечника. Гастроэнтерология. 2007. 133: 124–136. [PubMed] [Google Scholar] 94. Харрис Д.П., Хейнс Л., Сэйлс П.К., Дусо Д.К., Итон С.М., Лепак Н.М., Джонсон Л.Л., Суэйн С.Л., Лунд ИП. Взаимная регуляция продукции поляризованных цитокинов эффекторными В- и Т-клетками.Nat Immunol. 2000; 1: 475–482. [PubMed] [Google Scholar] 95. Мулен В., Андрис Ф., Тилеманс К., Малишевски С., Урбейн Дж., Мозер М. В-лимфоциты регулируют функцию дендритных клеток (ДК) in vivo: повышенное производство интерлейкина 12 ДК от В-клеток мышей с дефицитом приводит к отклонению типа 1 Т-хелперных клеток . J Exp Med. 2000; 192: 475–482. [Бесплатная статья PMC] [PubMed] [Google Scholar] 96. Bayry J, Lacroix-Desmazes S, Donkova-Petrini V, Carbonneil C, Misra N, Lepelletier Y, Delignat S, Varambally S, Oksenhendler E, Levy Y, Debre M, Kazatchkine MD, Hermine O, Kaveri SV.Природные антитела поддерживают дифференцировку и созревание дендритных клеток человека. Proc Natl Acad Sci U S. A. 2004; 101: 14210–14215. [Бесплатная статья PMC] [PubMed] [Google Scholar] 97. Радхакришнан С., Нгуен Л.Т., Чирик Б., Уре Д.Р., Чжоу Б., Тамада К., Донг Х., Ценг С.Ю., Шин Т., Пардолл Д.М., Чен Л., Кайл Р.А., Родригес М., Пиз Л.Р. Встречающиеся в природе антитела IgM человека, которые связывают B7-DC и усиливают стимуляцию Т-клеток дендритными клетками. J Immunol. 2003; 170: 1830–1838. [PubMed] [Google Scholar] 98. Амигорена С., Боннерот К.Рецепторы Fc для IgG и представление антигена на молекулах MHC класса I и класса II. Semin Immunol. 1999; 11: 385–390. [PubMed] [Google Scholar] 99. Игьетсеме Ю, Эко ФО, Хе Кью, Блэк СМ. Регулирование антителами Т-клеточного иммунитета: значение для стратегии вакцинации против внутриклеточных патогенов. Экспертные ревакцины. 2004; 3: 23–34. [PubMed] [Google Scholar] 100. Раветч Дж. В., Болланд С. Рецепторы IgG Fc. Анну Рев Иммунол. 2001; 19: 275–290. [PubMed] [Google Scholar] 101. Nimmerjahn F, Ravetch JV. Рецепторы Fcgamma: старые друзья и новые члены семьи.Иммунитет. 2006; 24: 19–28. [PubMed] [Google Scholar] 102. Десаи Д.Д., Харберс С.О., Флорес М., Колонна Л., Дауни М.П., Бергтольд А., Юнг С., Клайнс Р. Гамма-рецептор IIB Fc на дендритных клетках усиливает периферическую толерантность, ингибируя ответы эффекторных Т-клеток. J Immunol. 2007. 178: 6217–6226. [PubMed] [Google Scholar] 103. Bandukwala HS, Clay BS, Tong J, Mody PD, Cannon JL, Shilling RA, Verbeek JS, Weinstock JV, Solway J, Sperling AI. Передача сигналов через Fc {gamma} RIII требуется для оптимальных ответов Т-хелперов типа (Th) 2 и Th3-опосредованного воспаления дыхательных путей.J Exp Med. 2007; 204: 1875–1889. [Бесплатная статья PMC] [PubMed] [Google Scholar] 104. Nimmerjahn F, Ravetch JV. Дивергентная активность подкласса иммуноглобулинов g за счет селективного связывания рецептора Fc. Наука. 2005; 310: 1510–1512. [PubMed] [Google Scholar] 105. Ши Дж., Макинтош Р.С., Пасс Р.Дж. Терапевтические средства против малярии на основе антител и Fc-рецепторов. Clin Sci (Лондон) 2006; 110: 11–19. [PubMed] [Google Scholar] 106. Пак-Мин К.Х., Сербина Н.В., Янг В., Ма Х, Кристал Дж., Нил Б.Г., Натт С.Л., Ху Х, Ивашкив Л.Б. FcgammaRIII-зависимое ингибирование интерферон-гамма-ответов опосредует подавляющие эффекты внутривенного иммуноглобулина.Иммунитет. 2007; 26: 67–78. [PubMed] [Google Scholar] 107. Мур Т., Экворомаду СО, Эко Ф.О., Макмиллан Л., Рэйми К., Ананаба Г.А., Патриксон Дж. У., Нагаппан П.Р., Лин Д., Блэк С.М., Игьетсеме Ю. Опосредованная Fc рецептором регуляция антителами Т-клеточного иммунитета против внутриклеточных патогенов. J Infect Dis. 2003. 188: 617–624. [PubMed] [Google Scholar] 108. Йонето Т., Ваки С., Такай Т., Тагава Ю., Ивакура Ю., Мизугучи Дж., Нариучи Н., Йошимото Т. Критическая роль опосредованного рецептором Fc фагоцитоза, зависимого от антител, в устойчивости хозяина к инфекции XAT Plasmodium berghei на стадии крови.J Immunol. 2001; 166: 6236–6241. [PubMed] [Google Scholar] 109. Хубер В.К., Линч Дж. М., Бухер Д. Д., Ле Дж., Мецгер Д. В.. Фагоцитоз, опосредованный Fc-рецептором, вносит значительный вклад в избавление от инфекций вируса гриппа. J Immunol. 2001; 166: 7381–7388. [PubMed] [Google Scholar] 110. Kima PE, Constant SL, Hannum L, Colmenares M, Lee KS, Haberman AM, Shlomchik MJ, McMahon-Pratt D. Интернализация амастигот комплекса Leishmania mexicana через рецептор Fc необходима для поддержания инфекции при кожном лейшманиозе мышей.J Exp Med. 2000; 191: 1063–1068. [Бесплатная статья PMC] [PubMed] [Google Scholar] 111. Аппингтон Х., Менагер Н., Боросс П., Вуд Дж., Шеппард М., Вербеек С., Мастроени П. Влияние иммунной сыворотки и роль отдельных рецепторов Fcgamma на внутриклеточное распределение и выживаемость серовара Typhimurium Salmonella enterica в мышиных макрофагах. Иммунология. 2006. 119: 147–158. [Бесплатная статья PMC] [PubMed] [Google Scholar] 112. Menager N, Foster G, Ugrinovic S, Uppington H, Verbeek S, Mastroeni P. Рецепторы Fcgamma имеют решающее значение для выражения приобретенной устойчивости к вирулентному серовару Typhimurium Salmonella enterica in vivo, но не требуются для индукции гуморальных или Т-клеточных реакций. опосредованный иммунитет.Иммунология. 2007. 120: 424–432. [Бесплатная статья PMC] [PubMed] [Google Scholar] 113. Юань Р., Клайнс Р., О Дж., Раветч СП, Шарфф М.Д. Антитело-опосредованная модуляция инфекции Cryptococcus neoformans зависит от различных функций рецептора Fc и подклассов IgG. J Exp Med. 1998. 187: 641–648. [Бесплатная статья PMC] [PubMed] [Google Scholar] 114. Maglione PJ, Xu J, Casadevall A, Chan J. Гамма-рецепторы Fc регулируют иммунную активацию и восприимчивость во время инфекции Mycobacterium tuberculosis. J Immunol. 2008. 180: 3329–3338.[PubMed] [Google Scholar] 115. Климстра В.Б., Уильямс Дж.С., Райман К.Д., Хайднер Х.В. Нацеливание векторов на основе вируса Синдбис на Fc-рецептор-положительные типы клеток. Вирусология. 2005; 338: 9–21. [PubMed] [Google Scholar] 116. Тонгрен Дж. Э., Корран PH, Джарра В., Лангхорн Дж., Райли Е. М.. Эпитоп-специфическая регуляция переключения классов иммуноглобулинов у мышей, иммунизированных малярийными поверхностными белками мерозоитов. Infect Immun. 2005. 73: 8119–8129. [Бесплатная статья PMC] [PubMed] [Google Scholar] 117. Кальметт А. Пассивный иммунитет: попытки противотуберкулезной серотерапии.В: Calmette A, редактор. Туберкулезная палочка и туберкулез человека и животных. The Williams & Wilkins Co .; Балтимор, Мэриленд: 1923. С. 603–623. [Google Scholar] 118. Портер Р. Величайшая польза для человечества: история болезни человечества. W. W. Norton & Campany; Нью-Йорк, Нью-Йорк: 1997. [Google Scholar] 119. Глатман-Фридман А. Роль опосредованного антителами иммунитета в защите от Mycobacterium tuberculosis: продвижение к новой стратегии вакцинации. Туберкулез (Edinb) 2006; 86: 191–197.[PubMed] [Google Scholar] 120. Reljic R, Clark SO, Williams A, Falero-Diaz G, Singh M, Challacombe S, Marsh PD, Ivanyi J. Интраназальный IFN-гамма расширяет пассивную защиту мышей с помощью антител IgA против инфекции легких Mycobacterium tuberculosis. Clin Exp Immunol. 2006; 143: 467–473. [Бесплатная статья PMC] [PubMed] [Google Scholar] 121. Тейтельбаум Р., Глатман-Фридман А., Чен Б., Роббинс Дж. Б., Унануэ Е., Касадеваль А., Блум Б. MAb, распознающее поверхностный антиген Mycobacterium tuberculosis, увеличивает выживаемость хозяина.Proc Natl Acad Sci U S. A. 1998; 95: 15688–15693. [Бесплатная статья PMC] [PubMed] [Google Scholar] 122. Хамасур Б., Хайле М., Павловски А., Шредер Ю., Каллениус Г., Свенсон С.Б. Микобактериальные липоарабиноманнановые специфические моноклональные антитела и его F (ab ‘) фрагмент продлевают выживаемость мышей, инфицированных Mycobacterium tuberculosis. Clin Exp Immunol. 2004. 138: 30–38. [Бесплатная статья PMC] [PubMed] [Google Scholar] 123. Пете К., Алонсо С., Бьет Ф., Делогу Дж., Бреннан М.Дж., Лохт С., Меноцци Ф.Д. Гепарин-связывающий гемагглютинин M.tuberculosis требуется для внелегочного распространения. Природа. 2001; 412: 190–194. [PubMed] [Google Scholar] 124. Уильямс А., Релджик Р., Нейлор И., Кларк С.О., Фалеро-Диаз Г., Сингх М., Чалакомб С., Марш П.Д., Ивани Дж. Пассивная защита с помощью антител иммуноглобулина А против туберкулезной ранней инфекции легких. Иммунология. 2004. 111: 328–333. [Бесплатная статья PMC] [PubMed] [Google Scholar] 125. Хамасур Б., Хайле М., Павловски А., Шредер Ю., Уильямс А., Хэтч Г., Холл Г., Марш П., Каллениус Г., Свенсон С.Б.Конъюгаты арабиноманнана и белка Mycobacterium tuberculosis защищают от туберкулеза. Вакцина. 2003; 21: 4081–4093. [PubMed] [Google Scholar] 126. Фагарасан С., Хондзё Т. Т-независимый иммунный ответ: новые аспекты биологии В-клеток. Наука. 2000; 290: 89–92. [PubMed] [Google Scholar] 127. Боэс М. Роль естественных и иммунных антител IgM в иммунных ответах. Мол Иммунол. 2000; 37: 1141–1149. [PubMed] [Google Scholar] 128. Лопес-Карвалью Т., Кирни Дж. Ф. Развитие и отбор В-клеток маргинальной зоны.Immunol Rev.2004; 197: 192–205. [PubMed] [Google Scholar] 129. Аврора А.Б., Балук П., Чжан Д., Сидху С.С., Долганов Г.М., Басбаум С., Макдональд Д.М., Киллин Н. Зависимое от иммунного комплекса ремоделирование сосудистой сети дыхательных путей в ответ на хроническую бактериальную инфекцию. J Immunol. 2005; 175: 6319–6326. [PubMed] [Google Scholar] 130. Клайнс Р. Защитные механизмы ВВИГ. Curr Opin Immunol. 2007. 19: 646–651. [PubMed] [Google Scholar] 131. Андерсен П. Противотуберкулезные вакцины — обновленная информация. Nat Rev Microbiol. 2007. 5: 484–487.[PubMed] [Google Scholar] 132. Фаучи А.С., Джонстон М.И., Диффенбах С.В., Бертон Д.Р., Хаммер С.М., Хокси Д.А., Мартин М., Овербо Дж., Уоткинс Д.И., Махмуд А., Грин В. Исследования вакцины против ВИЧ: путь вперед. Наука. 2008. 321: 530–532. [PubMed] [Google Scholar] 133. Мур К.В., де Ваал Малефит Р., Коффман Р.Л., О’Гарра А. Интерлейкин-10 и рецептор интерлейкина-10. Анну Рев Иммунол. 2001; 19: 683–765. [PubMed] [Google Scholar] 134. Жункейра-Кипнис А.П., Кипнис А., Энао Тамайо М., Хартон М., Гонсалес Хуарреро М., Басараба Р.Дж., Орме И.М.Продукция интерлейкина-10 макрофагами легких у мышей с мутантом CBA xid, инфицированных Mycobacterium tuberculosis. Иммунология. 2005. 115: 246–252. [Бесплатная статья PMC] [PubMed] [Google Scholar] 135. Каманака М., Ким С.Т., Ван Й.Й., Сатервала Ф.С., Лара-Техеро М., Галан Дж. Э., Хархадж Е., Флавелл Р.А. Экспрессия интерлейкина-10 в лимфоцитах кишечника, обнаруженная тигровой мышью, нокаутирующей репортер интерлейкина-10. Иммунитет. 2006; 25: 941–952. [PubMed] [Google Scholar] 136. Мейнард К.Л., Харрингтон Л.Е., Яновски К.М., Оливер Дж. Р., Зиндл К.Л., Руденский А.Ю., Уивер К.Т.Регуляторные Т-клетки, экспрессирующие интерлейкин 10, развиваются из клеток-предшественников Foxp3 (+) и Foxp3 (-) в отсутствие интерлейкина 10. Nat Immunol. 2007; 8: 931–941. [PubMed] [Google Scholar] 137. Рутелла С., Данезе С., Леоне Г. Толерогенные дендритные клетки: модуляция цитокинов достигает зрелости. Кровь. 2006; 108: 1435–1440. [PubMed] [Google Scholar] 138. Olson TS, Bamias G, Naganuma M, Rivera-Nieves J, Burcin TL, Ross W., Morris MA, Pizarro TT, Ernst PB, Cominelli F, Ley K. Расширенная популяция В-клеток блокирует регуляторные Т-клетки и обостряет илеит на мышиной модели болезни Крона.J Clin Invest. 2004. 114: 389–398. [Бесплатная статья PMC] [PubMed] [Google Scholar] 139. Mizoguchi A, Bhan AK. Случай для регуляторных В-клеток. J Immunol. 2006. 176: 705–710. [PubMed] [Google Scholar] 140. Чжун Г., Рейс и Соуза С., Жермен Р.Н. Как антиген-неспецифические В-клетки, так и лимфоидные дендритные клетки демонстрируют обширную поверхностную экспрессию процессированных комплексов класса II главного комплекса гистосовместимости после воздействия растворимого белка in vivo или in vitro. J Exp Med. 1997; 186: 673–682. [Бесплатная статья PMC] [PubMed] [Google Scholar] 141.Грей Д., Грей М., Барр Т. Врожденные ответы В-клеток. Eur J Immunol. 2007. 37: 3304–3310. [PubMed] [Google Scholar] 143. Дхеда К., Бут Х, Хаггетт Дж. Ф., Джонсон М. А., Зумла А., Ладья Г. А.. Ремоделирование легких при туберкулезе легких. J Infect Dis. 2005; 192: 1201–1209. [PubMed] [Google Scholar] 144. Мораес Т.Дж., Зуравска Дж.Х., Дауни Г.П. Содержание нейтрофильных гранул в патогенезе поражения легких. Curr Opin Hematol. 2006; 13: 21–27. [PubMed] [Google Scholar] 145. Ерусланов Е.Б., Лядова И.В., Кондратьева Т.К., Майоров КБ, Щеглов И.В., Орлова М.О., кв.Ответ нейтрофилов на инфекцию Mycobacterium tuberculosis у генетически восприимчивых и устойчивых мышей. Infect Immun. 2005; 73: 1744–1753. [Бесплатная статья PMC] [PubMed] [Google Scholar]Os gringos nos amam | Эдублин
É incrível! Eles ficam felizes quando descobrem de onde somos. Фотография: Shutterstock
.Coisa mais legal de morar no external é dizer para os gringos que vê é brasileiro. Desde que cheguei aqui venho colecionando reações das mais Variadas. Ума вез, ум irlandês duvidou da minha resposta por eu ter olhos verdes.Tive que «provar».
— Итак, говори по-португальски, чувак!
— Isso é sério ou é zoeira?
— Я знаю «зоэйру»! Ты паулиста, велхо!
Amigos nisseis e sanseis Precisam fazer o mesmo quando encontram asiáticos. Assim como eu, um colega também foi desafiado para um duelo de idiomas.
— Ваташи ва аната га бураджирухито нару то ва кангэте инаи.
— Эй?
A noiva de um brother teve que explicar na imigração como tinha passaporte espanhol se era brasileira.Ela até pensou em tirar onda com a cara da oficial. «Comprei no Classificados Dublin. Já ouviu falar? » . Mas para evitar issues, acabou contando a história verdadeira. Achei prudente da parte dela.
Certa vez, após me perguntar de onde eu era, um inglês desandou a fazer perguntas sobre violência. Queria saber se era verdade que estávamos sequestrando europeus durante a Copa do Mundo.
— Кларо! Depois vamos pedir resgate pro Governos deles.
— Правда?
— Ахам.Quanto a rainha pagaria por vêê?
Ele ficou assustado, eu dee que era brincadeira e fui obrigado a explicar que apenas brasileiros são autorizados a falar mal do Brasil.
— Eu Posso! Você não!
Então Passi a contar-lhe histórias sobre os adoráveis seres que desiream o nosso Congresso Nacional.
Um italiano nem deixou eu terminar de Responder e já disparou a falar sobre futebol. Belga e francesa começaram с песней Мишеля Тело. O alemão quis saber mais sobre o nosso «espírito festeiro».A polonesa ficou toda feliz por encontrar um latino para praticar espanhol.
— Puede ser portuñol?
É incrível! Eles ficam felizes quando descobrem de onde somos. E o mais interessante é que eles encontram brasileiros TODOS OS DIAS. Não deveria ser tão novidade assim…
— Por que tem tanta gente do Brasil vindo para cá?
— O que acontece no país de voiceês que todo mundo tá fugindo de lá?
— Tem mais gente vindo? Acho que não tem mais espaço.
— Vocês estão colizando a Irlanda?
Сим, эстамос! Na verdade, tudo faz parte de um plano maligno do Governo brasileiro para exportar nossa cultura mundo afora, começando pelo povo Irish. Нет окончательного отношения enviado в terra mãe pelos bravos soldados que aqui estão, возможно, проверенных конкистами, которые считают. De acordo com o documento sigiloso, 100% dos brasileiros residences em Dublin estavam ou estão ensinando português para pelo menos um gringo. Eu faço a minha parte.Французский já sabe mandar os caras mais assanhados da balada tomar naquele lugar. A irlandesa do trabalho nunca mais falou «доброе утро». Ela acha «bom dia» muito mais legal e animado. О espanhol semper pergunta se eu quero «breja». A americana só agradece com «obrigada». A polonesa que queria falar espanhol virou minha namorada e vive pedindo «carinho». O coreano, surpreendentemente mais atrasado do que os demais no aprendizado, ainda está na fase do «nossa, nossa, assim voiceê me mata».
Confesso que sinto-me extremamente bem quando vejo um estrangeiro tentando aprender a língua portuguesa.Viemos para cá estudar a língua dos caras e agora são os caras que querem estudar a nossa língua. Сюрреалистично! E mais do que isso, eles parecem desejar ter o mesmo estilo de vida que o nosso: abraços, risos, farra, calor humano, churrasco, cerveja gelada. Ничего подобного юмора нет.
Obviamente, essa relação globalizada é carregada de estereótipos, o que não é needariamente ruim. Afinal, são justamente os pré-Conceitos que despertam a curiosidade e os fazem se aprofundar mais em nossa cultura.Aí, cla, depois de um tempo em nosso meio, eles percebem que nem semper estamos em festa (com exceção dos vizinhos brasileiros do andar de cima). Quando o nível de intimidade chega neste ponto, já é tarde demais: voiceê e o gringo já se tornaram grandes amigos.
Eis que wave o maior проблема do mundo para todo brasileiro que mora no external: onde Hospedar todos os amigos estrangeiros quando eles forem te visitar no paísropical, abençoado por Deus e bonito por natureza? Pelas minhas contas, Precisaria reservar um hotel inteiro para cumprir as promessas de hospitalidade já feitas.Ou talvez eu Possa distribuí-los nas casas da irmã, do pai, da mãe e de alguns outros parentes. E depois disso? O que fazer? Eles já me disram querem querem conhecer Rio de Janeiro, Nordeste (inteiro), Floripa e a Floresta Amazônica. «É tudo muito longe», реклама. «Não se preocupe, a gente aluga um carro», — responsedeu o animado coreano. Dei risada. Ele também, embora não tenha entendido o motivo de tanta graça.
Na semana passada eu chamei seis gringos para irem ao pub assistir um jogo do Brasil.Eles aparecerem com camisas da seleção. Todos. Torceram como nunca. Беррарам, гритарам, беберам, селеберарам. Mas o sorriso estampado no rosto do coreano sumiu quando um colega puxou assunto.
— Ты откуда?
— Корея.
Ele queria poder dizer «Бразилия».
Revisado por Tarcísio Junior
Изображения через Shutterstock
Encontrou algum erro or quer nos comunicar uma informação?
Envie uma mensagem para jornalismo @ edublin.com.br
Краткий справочник букв столбцов Excel для номеров
Вот краткий справочник по сопоставлению букв столбцов Excel с номерами. Много раз мне нужно было найти номер столбца, связанный с буквой столбца, чтобы использовать его в макросе Excel. Для такого ленивого разработчика, как я, использование моих математических навыков для получения ответа отнимает очень много времени, поэтому я создал этот быстрый справочник для себя.
Перейти к определенному списку столбцов
Если вы ищете конкретный столбец, щелкните следующие ссылки, чтобы перейти к определенному списку столбцов
Столбцы Excel AZ
| Буква столбца | Номер столбца |
|---|---|
| A | 1 |
| B | 2 |
| C | 3 |
| D | 4 |
| E | G | 7 |
| H | 8 |
| I | 9 |
| J | 10 |
| K | 11 | 13 |
| N | 14 |
| O | 15 |
| P | 16 |
| Q | 17 |
| R | 18 |
| S | 19 |
| T | 20 |
| U | 90521 | 905W | 23 |
| X | 24 |
| Y | 25 |
| Z | 26 |
Колонки Excel
Столбцы Excel 9050
Колонки Excel CA-CZ
| 79 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CB | 80 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CC | 81 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CD | 82 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CE | 83 | 905 905 905 905 905 905 CF 905 85|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CH | 86 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CI | 87 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CJ | 88 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CK | 89 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CN | 92 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CO | 93 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CP | 9 4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CQ | 95 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CR | 96 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CS | 97 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CT | 98 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CW | 101 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CX | 102 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CY | 103 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CZ | 104 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DA | 105 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DB | 106 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DC | 107 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DD | 108 | 905DD | 108 | 109511110 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DG | 111 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DH | 112 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DI | 113 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DJ | 114 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DK | 115 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DL | 116 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 117 DM | 5 DO119 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DP | 120 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DQ | 121 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DR | 122 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DS 9015 | 123 | 905 905 905 905 905 905 905 905 905 905 905125 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DV | 126 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DW | 127 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DX | 128 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DY | 129 |
| Буква столбца | Номер столбца | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 131 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EB | 132 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EC | 133 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ED | 134 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EE | 135 905 905 905 EE | 135 905 | 137 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EH | 138 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EI | 139 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EJ | 140 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EK | 141 905 905 905 905 905 905 EL 905 905 905 EL 905 143 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EN | 144 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EO | 145 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EP | 146 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EQ | 147 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ET | 150 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EU | 151 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EV | 9 0515152||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EW | 153 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EX | 154 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EY | 155 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EZ | 156 | EZ | 156 | 16Номер столбца | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FA | 157 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FB | 158 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FC | 159 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FF | 162 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FG | 163 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FH | 164 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FI | 165 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FJ 166 905 905 905 905 905 905 905 905 905 905 905 905 905 905 905 | 168 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FM | 169 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FN | 170 90 516 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FO | 171 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FP | 172 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FQ | 173 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FR | 174 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FU | 177 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FV | 178 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FW | 179 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FX | 180 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Буква столбца | Номер столбца |
|---|---|
| GA | 183 |
| GB | 184 |
| GE | 187 |
| GF | 188 |
| GG | 189 |
| GH | 190 |
| GI | 191 |
| GJ | 192 |
| 195 | |
| GN | 196 |
| GO | 197 |
| GP | 198 |
| GQ | 199 | 905GQ | 199 | 905201 |
| GT | 202 |
| GU | 203 |
| GV | 204 |
| GW | 205 | 905 905 905 905 905
| GZ | 208 |
Столбцы Excel HA-HZ
| Буква столбца | Номер столбца |
|---|---|
| HA | 209 |
| HB | 210 |
| HC | 211 | 905 905 905 905 905 905 HE 15
| HF | 214 |
| HG | 215 |
| HH | 216 |
| HI | 217 | 905 905 905 215
| HL | 220 |
| HM | 221 |
| HN | 222 |
| HO | 905 223 |
| 905 905 905 223 | |
| HP 905 HR | 226 |
| HS | 227 |
| HT | 228 |
| HU | 229 |
| HV | 230 |
| HW | 231 |
| HX | 232 905 905 905 |
Столбцы Excel IA-IZ
| Буква столбца | Номер столбца | |
|---|---|---|
| IA | 235 | |
| IB | 905 905 905ID | 238 |
| IE | 239 | |
| IF | 240 | |
| IG | 241 | |
| IH | 905 905 905 905 905 905 905 905 IIH 905 905 II244 | |
| IK | 245 | |
| IL | 246 9 0516 | |
| IM | 247 | |
| IN | 248 | |
| IO | 249 | |
| IP | 250 | 905 905 905 905 | 905 905 905 IQ
| IS | 253 | |
| IT | 254 | |
| IU | 255 | |
| IV | 256 | 259 |
| IZ | 260 |
Колонки Excel JA-JZ
| Колонка Letter | Номер колонки |
|---|---|
| JC | 263 |
| JD | 264 |
| JE | 265 |
| JF | 266 |
| JG | 267 |
| JH | 268 | 271 |
| JL | 272 |
| JM | 273 |
| JN | 274 |
| JO 905 905 905 905 905 905 905 905 JO 905 | 277 |
| JR | 278 |
| JS | 279 |
| JT | 280 |
| JU | 281 905 905 905 905 905 |
| JX | 284 |
| JY | 285 |
| 286 |
Столбцы Excel KA-KZ
| Буквенное обозначение столбца | Номер столбца |
|---|---|
| KA | 287 | 289 |
| KD | 290 |
| KE | 291 |
| KF | 292 |
| KG | 293 905 905 |
| KJ | 296 |
| KK | 297 |
| KL | 298 |
| KM | 299 | 905 905 905 905 905 905 15 299 905 905 905 905 905 299 905
| KP | 302 |
| KQ | 303 |
| KR | 304 |
| KS | 305 |
| KT | 306 |
| KU | 307 |
| KV | 308 905 905 905 905 |
| KY | 311 |
| KZ | 312 |
Столбцы Excel LA-LZ
| Буква столбца | LB | 314 |
|---|---|
| LC | 315 |
| LD | 316 |
| LE | 317 |
| LF | 905 905 905 317 905320 |
| LI | 321 |
| LJ | 322 |
| LK | 323 |
| LL | 324 |
| LM | 325 |
| LN | 326 |
| LQ | 329 |
| LR | 330 |
| LS | 331 |
| LT | 332 |
| 3315 905 905 905 905 905 LW | 335 |
| LX | 336 |
| LY | 337 |
| LZ | 338 |
Excel 907 Колонки 905 905 9050 9050 M-905
Столбцы Excel NA-NZ
| Колонка 905 905 905 905 905 905 905 905 905 905 Колонка 905 Буква | 365 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NB | 366 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NC | 367 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ND | 368 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NE | 368 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NE | 369 | 905 905 905|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NH | 372 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NI | 373 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NJ | 374 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NK | 375 | 905 905 905 905 905 905 905 NK 905|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NN | 378 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NO | 379 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NP | 380 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NQ | 381 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NR | 382 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NS | 383 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NT | NV384 905 905 905 905 905 905 905 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NW | 387 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NX | 388 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NY | 389 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NZ | 390 | 905 905 Letter-9050 9050 905 905 905 905 905 905 905 Letter-905Номер столбца | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OA | 391 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OB | 392 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OC | 393 | OD16 905 905 905 905 905 905 OF5 | 396 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OG | 397 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OH | 398 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OI | 399 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OJ | 400 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ОК | 401 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OL | 402 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OO | 405 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OP | 406 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OQ | 407 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OR | 408 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 905 905 905 905 905 905 905 905 905 905 OS 905 OU | 411 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OV | 412 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OW | 413 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OX | 414 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
OY1615 905 905 905 905 905 905 905 414 905 905 Столбцы PA-PZ
Столбцы Excel RA-RZ
Столбцы Excel SA-SZ 9017 8
Столбцы Excel TA-TZ
Столбцы Excel UA-UZ
Столбцы Excel VA-VZ
Excel Columns WA-WZ
Excel Columns XA-XZ
Excel Columns YA-YZ
Excel Columns ZA-ZZ
DT Neville Gallimore is primed to fill a big void in the Cowboys defense in 2021The defensive tackle position for the Cowboys has been a major issue for some time now.Как и позиция безопасности, это позиция защиты Cowboys, в которую до недавнего времени главный офис не проявлял особого интереса к инвестированию ресурсов. Невилл Галлимор, выбранный в третьем раунде 2020 года, был одним из самых талантливых игроков, которых Ковбои приобрели на этой позиции. Другие попытки до этого года включали подписание в свободном агентстве стареющего ветерана Джеральда Маккоя, который проиграл из-за травмы в конце сезона в тренировочном лагере, и выбор во втором раунде Тристен Хилла, который пока не удался. Галлимор вошел в драфт НФЛ 2020 года с некоторым шумом в первом раунде, но шумиха в конце концов утихла, чем ближе мы подошли к драфту. Тем не менее, он услышал, как его имя назвали 82-м номером в общем пике, и оказался в битве за игровое время после прыжка в Далласе в качестве новичка. В его сезоне новичка были свои взлеты и падения, но, заработав стартовую роль из-за недостатка таланта и травм в обороне, Галлимор действительно оказался на финишной прямой в сезоне.Он закончил свой сезон новичков с пол-мешка, но закончил последние шесть недель сезона с двумя спешками, тремя ударами QB и шестью прессингами. Первые сообщения из Окснарда гласят, что Галлимор действительно хорошо выглядит. Согласно многочисленным сообщениям, Галлимор выглядит больше, более взрывоопасен и готов совершить большой второй год прыжка за Ковбоев на позиции защитного захвата. В начале сезона таблица глубины DT, вероятно, была одной из самых слабых групп в списке.Поскольку Галлимор, как ожидается, выиграет одну из стартовых позиций, ветераны Карлос Уоткинс и Брент Урбан, вероятно, будут бороться за второе место в стартовом защитном отборе вместе с Невиллом. Помимо этой большой тройки, Ковбои будут полагаться на новичков Осу Одигизуву, Куинтона Боханну и, возможно, Тристен Хилла, в зависимости от того, когда он сможет поработать после реабилитации ACL, которую он сейчас проводит в лагере. Без тонны опыта в нижней части диаграммы глубины Ковбои будут в значительной степени полагаться на Невилла Галлимора, который сыграет для них огромную роль в 2021 году, и пока что кажется, что он справится с этой задачей. |