В чем разница между государственным пенсионным фондом и негосударственным?
Как выбрать управляющую компанию или негосударственный пенсионный фонд для формирования накопительной пенсии?
В чем разница между государственным пенсионным фондом и негосударственным? М. Монгуш, Кызыл
Отвечает руководитель группы по инвестированию пенсионных накоплений ОПФР по Республике Тыва Чимис Эрес-ооловна Монгуш.
Вы можете спросить «Зачем мне слушать про пенсию сейчас, когда мне всего 20,30 или 40лет и до нее еще, ох, как далеко?!» Но не торопитесь отмахиваться от этой темы. Это здорово, что вам до пенсии пока далеко! Ведь, чем раньше начнете задумываться о будущей пенсии, тем больше успеете сделать, чтобы увеличить ее размер.
- Для этого необходимо контролировать платит ли работодатель взносы за Вас в ПФР.
- Грамотно управлять своими пенсионными накоплениями.

Система обязательного пенсионного страхования (ОПС) действует в России с 2002 года и базируется на страховых принципах: основой будущей пенсии гражданина являются страховые взносы, которые уплачивают за него работодатели в течение всей трудовой жизни.
Чтобы стать участником системы ОПС и формировать свои пенсионные права, нужно быть зарегистрированным в системе индивидуального (персонифицированного) учета ПФР. В этой системе в течение всей трудовой деятельности гражданина фиксируются данные, необходимые для назначения, выплаты и перерасчета пенсии: о стаже, периодах трудовой деятельности и местах работы, и в первую очередь – о страховых взносах, поступивших в фонд его будущей пенсии.
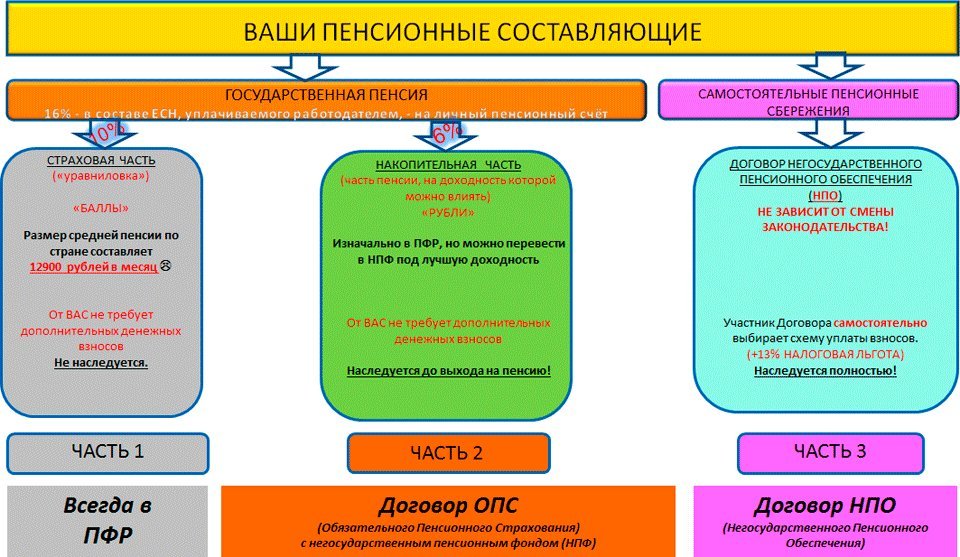
Страховые взносы в ОПС работодатели платят по тарифу 22% от фонда оплаты труда работника. Из них 6% тарифа могут идти на формирование пенсионных накоплений, а 16% – на формирование страховой пенсии, а могут, по выбору гражданина, все 22% идти на формирование страховой пенсии.
Важно знать! В 2014 — 2019 годах по решению государства все средства страховых взносов на обязательное пенсионное страхование, уплачиваемые работодателями за своих работников, направляются на формирование страховой пенсии, т. е. все 22% идут на формирование только страховой пенсии.
е. все 22% идут на формирование только страховой пенсии.
Накопительная часть будущей пенсии формируется у работающих граждан 1967 года рождения и моложе, а также у участников Программы государственного софинансирования пенсий, уплачивающих дополнительные страховые взносы.
Граждане, у которых формируются средства пенсионных накоплений в системе ОПС, вправе передавать эти средства страховщику для инвестирования на финансовом рынке государственной или частными управляющими компаниями. Страховщиком пенсионных накоплений может быть Пенсионный фонд России или негосударственный пенсионный фонд, входящий в систему гарантирования прав граждан. ПФР инвестирует пенсионные накопления через государственную управляющую компанию «Внешэкономбанк» и частные управляющие компании.
В Российской Федерации есть два типа пенсионных фондов — государственный (ПФР) и негосударственные (НПФ).
Государственный пенсионный фонд
В России на данный момент работает только один государственный пенсионный фонд — Пенсионный фонд Российской Федерации (ПФР).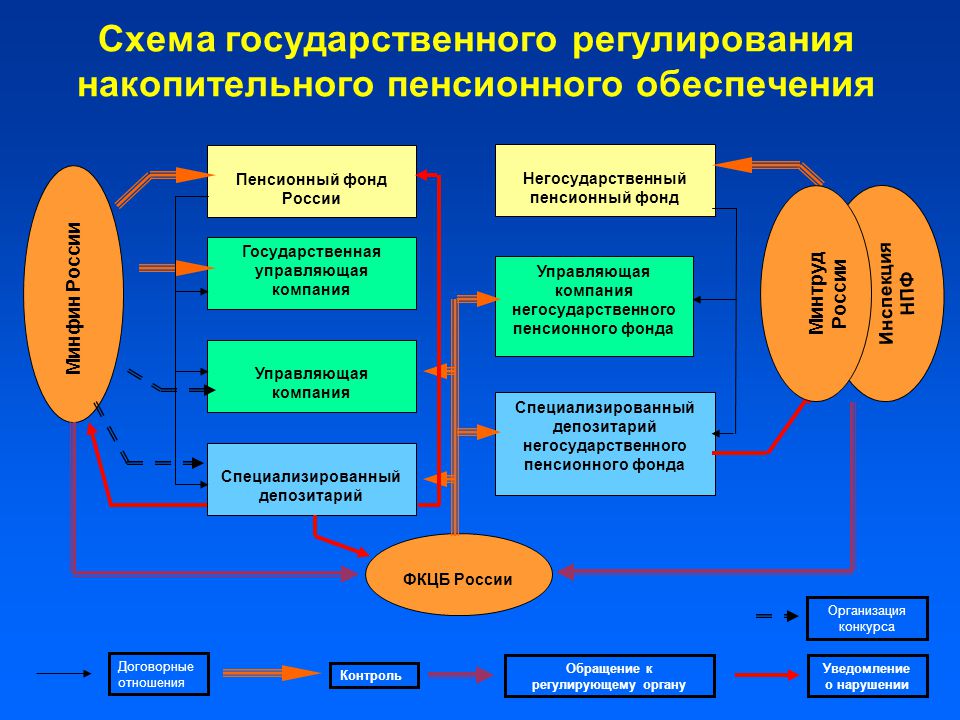 ПФР является крупнейшей федеральной системой оказания государственных услуг в области социального обеспечения в России. Пенсионный фонд России входит в бюджетную систему страны и выполняет широкий круг социально значимых задач, связанных с пенсионным обеспечением и пенсионным страхованием граждан РФ.
ПФР является крупнейшей федеральной системой оказания государственных услуг в области социального обеспечения в России. Пенсионный фонд России входит в бюджетную систему страны и выполняет широкий круг социально значимых задач, связанных с пенсионным обеспечением и пенсионным страхованием граждан РФ.
Негосударственные пенсионные фонды (НПФ)
Негосударственный пенсионный фонд – некоммерческая организация социального обеспечения. На рынке пенсионного страхования НПФ осуществляют несколько видов деятельности, в том числе деятельность по негосударственному пенсионному обеспечению участников НПФ в соответствии с договорами негосударственного пенсионного обеспечения. НПФ, как и ПФР, может выступать страховщиком гражданина в части его пенсионных накоплений. По достижении человеком пенсионного возраста НПФ обязан выплачивать накопительные пенсии, исходя из размера накоплений гражданина, сформированных на его счете в НПФ.
В чем отличие государственного пенсионного фонда от негосударственного
Главное отличие государственного пенсионного фонда от негосударственного в том, что ПФР включен в бюджетную систему и полностью подотчетен государству, а НПФ —не входит в бюджетную систему РФ и является частной компанией.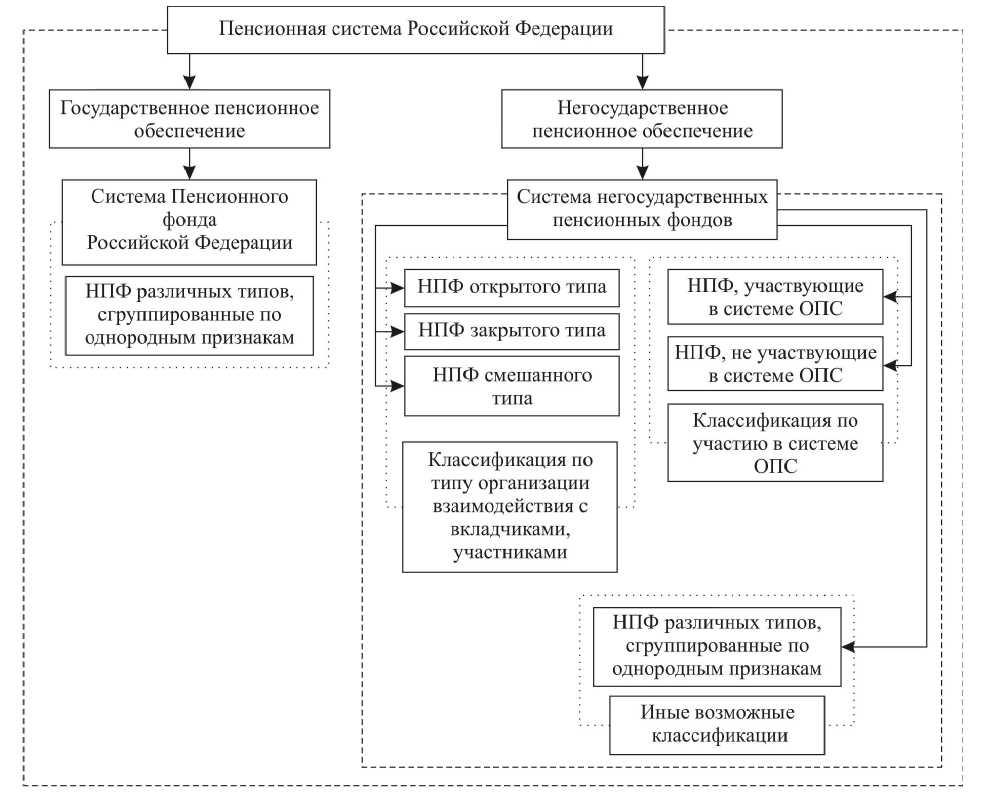
Выбор НПФ только за Вами
Необходимо внимательно подходить к выбору негосударственного пенсионного фонда (НПФ), принимать во внимание опыт, репутацию НПФ и показатели доходности от инвестирования пенсионных накоплений. Полная информация о негосударственных пенсионных фондах размещена на сайте Банка России и на сайтах самих НПФ. Список негосударственных пенсионных фондов, которые вошли в систему гарантирования прав застрахованных лиц, размещен на сайте Банка России и Агентства по страхованию вкладов.
Выбор страховщика – это личное решение гражданина. Требовать перевода средств пенсионных накоплений в НПФ не имеют права ни работодатель, ни агентства по трудоустройству, ни коммерческие банки при кредитовании.
Варианты подачи заявлений о переводе пенсионных накоплений в УК и НПФ:
Через клиентскую службу ПФР или МФЦ
- Заявление можно подать как лично, так и через представителя.

Через интернет
- Подать электронное заявление можно через Единый портал государственных услуг или Личный кабинет гражданина на сайте ПФР. И в том, и в другом случае электронное заявление должно быть подписано усиленной квалифицированной электронной подписью.
По почте
- В этом случае установление личности гражданина и проверка подлинности его подписи осуществляется нотариусом или в установленном законодательстве порядке.
СОВЕТ!
Если Вы принимаете решение о смене страховщика, то есть о переводе средств пенсионных накоплений из ПФР в НПФ или из НПФ в ПФР, чтобы избежать потери инвестиционного дохода или убытков от инвестирования пенсионных накоплений, рекомендуется подавать заявление о переходе к новому страховщику через пять лет. Менять страховщика, переводя свои пенсионные накопления чаще одного раза в пять лет, не выгодно.
В случае возникновения дополнительных вопросов Вы можете обратиться в Отделение Пенсионного фонда Российской Федерации по Республике Тыва по адресу: г.Кызыл, ул.Кочетова, д.18, каб. 308, тел. (394 22) 9-61-20.
Поделиться новостью
Что делать, если пенсионные накопления переведены без согласия гражданина?
Проблема в нашей стране неправомерного перевода средств пенсионных накоплений в негосударственные пенсионные фонды существует достаточно давно, с момента выхода негосударственных пенсионных фондов (далее — НПФ) на рынок предоставления услуг по обязательному пенсионному страхованию.
Практика неправомерного перевода средств пенсионных накоплений показывает, что недобросовестные агенты негосударственных пенсионных фондов используют неправомерные способы — без ведома граждан, но от их имени — оформления договоров об обязательном пенсионном страховании и заявлений о досрочном переходе в НПФ.
На законодательном уровне регулярно вносятся нормы ужесточающие условия участия негосударственных пенсионных фондов в обязательном пенсионном страховании. Так, в 2014 году Банком России проведена проверка НПФ на финансовую устойчивость, отменена подача заявлений через негосударственные пенсионные фонды и кредитные организации на основании заключенных соглашений о взаимном удостоверении подписей. В середине 2017 года отменена подача заявлений в электронной форме через удостоверяющие центры.
Теперь для подачи заявлений о переходе или о досрочном переходе в негосударственный пенсионный фонд граждане лично или через представителя по доверенности обращаются в любой территориальный орган ПФР или направляют заявления через Единый портал государственных услуг при наличии усиленной квалифицированной электронной подписи.
Как избежать неправомерного перевода накоплений?
Нередко ситуация с неправомерным переводом средств происходит по причине необдуманного распространения гражданами своих персональных данных. Поскольку для оформления с негосударственным пенсионным фондом договора об обязательном пенсионном страховании и оформлении заявления, на основании которых осуществляется перевод средств, необходимо знать только страховой номер индивидуального лицевого счета (СНИЛС) и Ф.И.О. гражданина.
В связи с этим рекомендуем гражданам отслеживать информацию о действующем страховщике по обязательному пенсионному страхованию на портале госуслуг РФ в разделе «Пенсионные накопления» по адресу: http://www.gosuslugi.ru/ и на официальном сайте ПФР в Личном кабинете гражданина: www.pfr.gov.ru или при личном обращении в клиентскую службу территориального органа ПФР.
Как вернуть накопления, оказавшиеся без Вашего заявления в чужом негосударственном пенсионном фонде?
В случае выявления факта неправомерного перевода средств, гражданину следует направить письменное обращение в территориальный орган ПФР или в электронной форме на сайте ПФР ( www.
Обращаем внимание, что неправомерно заключенный договор с негосударственным пенсионным фондом можно признать недействительным только в судебном порядке. Это единственный способ вернуть пенсионные накопления к прежнему страховщику и возместить потери инвестиционного дохода. Так согласно п. 5.3 ст. 36.6 Федерального закона от 7 мая 1998 г. №75-ФЗ «О негосударственных пенсионных фондах» при наличии у гражданина решения суда о признании договора недействительным, негосударственный пенсионный фонд в течение 30 дней обязан перевести предыдущему пенсионному фонду полученные средства пенсионных накоплений, инвестиционный доход, не учтенный на момент перевода ему указанных средств, а также проценты за неправомерное пользование средствами пенсионных накоплений, определяемые в соответствии со ст. 395 ГК РФ.
395 ГК РФ.
ПФР или НПФ: в чем разница между ними, плюсы и минусы
Всем жителям России уже почти два десятка лет предоставлен выбор, где формировать накопительную пенсию – в Пенсионном фонде России (ПФР) или негосударственных пенсионных фондах (НПФ). Если деньги в ПФР, то ими управляет ВЭБ.РФ или частные управляющие компании, с которыми заключил договор Пенсионный фонд России. А инвестированием средств, размещенных в НПФ, занимаются те частные управляющие компании, с которыми заключил договор НПФ. Для того чтобы выбрать, кому доверить свои накопления, нужно взвесить все плюсы и минусы каждого типа организации.
Как формируются пенсионные накопления?Наша пенсия делится на две части – страховую и накопительную. До 2014 года страховую часть пенсии – 16% от дохода сотрудников – работодатель в обязательном порядке перечислял в ПФР на выплату пенсий нынешним пенсионерам, а еще 6% перечислял на личный счет работника в ПФР, или НПФ. Эти деньги (6%) до сих пор находятся на индивидуальном лицевом счете гражданина и будут выплачиваться ему после выхода на пенсию. На отчисления в накопительную часть пенсии действует мораторий с 2014 по 2021 год, в этот период все 22% отчислений идут на страховую пенсию.
Эти деньги (6%) до сих пор находятся на индивидуальном лицевом счете гражданина и будут выплачиваться ему после выхода на пенсию. На отчисления в накопительную часть пенсии действует мораторий с 2014 по 2021 год, в этот период все 22% отчислений идут на страховую пенсию.
Подробнее – читайте статью «Заморозка пенсионных накоплений: что это значит?».
Куда вкладывают деньги управляющие компании ПФР и НПФ?Накопительная пенсия россиян может храниться как в ПФР, так и в НПФ. Основная задача страховщиков – сберечь накопления и обеспечить их доходность на уровне не ниже инфляции (как минимум). Сверхзадача – обеспечить серьезную прибавку к будущей пенсии за счет высокого дохода от инвестирования. Для этого управляющие компании ПФР и НПФ инвестируют сбережения в различные финансовые инструменты: облигации, государственные ценные бумаги, депозиты и так далее. Важно: сами фонды в настоящий момент не инвестируют пенсионные накопления. Эту работу по договору с ними ведут управляющие компании. Это называется «доверительное управление».
Это называется «доверительное управление».
«Стиль» инвестирования – одно из главных отличий государственного пенсионного фонда от частного. По умолчанию ПФР передает средства в управление государственной управляющей компании (ГУК) ВЭБ.РФ. ГУК ВЭБ.РФ придерживается консервативной инвестиционной политики, инвестируя только в депозиты, облигации и госбумаги. Возможности НПФ в целом шире: они имеют право до 10% от средств вкладывать в высокорискованные активы (например, акции). Такие инвестиции сулят большую прибыль, но также могут принести и убытки, если, например, котировки акций пойдут вниз. По этой же причине в рейтингах доходности НПФ могут занимать как первые, так и последние места по доходности: многое зависит от того, угадал ли фонд с выбором высокорискованных активов. А ГУК ВЭБ.РФ, несмотря на консервативный характер инвестирования, находится, как правило, среди «крепких середняков»: доходности его портфелей превышают инфляцию и результаты ряда НПФ.
Как защищены средства?В отличие от частных фондов ни ПФР, ни ГУК ВЭБ.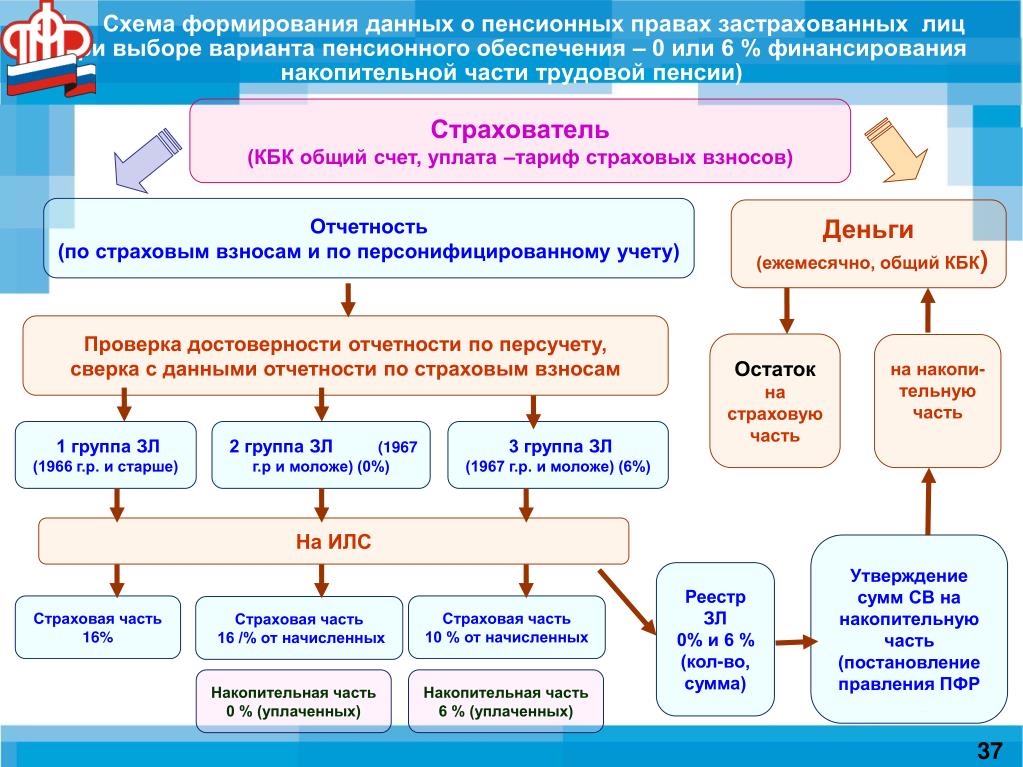 РФ не могут лишиться лицензии или обанкротиться. Однако банкротство или отзыв лицензии у НПФ (такое иногда случается) хоть и неприятны, но некритичны, ведь все накопления в системе обязательного пенсионного страхования (ОПС) гарантированы государством. И если у НПФ отзовут лицензию или он объявит себя банкротом, граждане, которые держали в нем деньги, не лишатся накоплений. Все сбережения застрахованы Агентством по страхованию вкладов (вот тут можно посмотреть список НПФ – участников системы страхования), и в случае проблем у НПФ они будут переведены в ПФР. Однако инвестиционный доход за последние несколько лет при таком сценарии может быть потерян.
РФ не могут лишиться лицензии или обанкротиться. Однако банкротство или отзыв лицензии у НПФ (такое иногда случается) хоть и неприятны, но некритичны, ведь все накопления в системе обязательного пенсионного страхования (ОПС) гарантированы государством. И если у НПФ отзовут лицензию или он объявит себя банкротом, граждане, которые держали в нем деньги, не лишатся накоплений. Все сбережения застрахованы Агентством по страхованию вкладов (вот тут можно посмотреть список НПФ – участников системы страхования), и в случае проблем у НПФ они будут переведены в ПФР. Однако инвестиционный доход за последние несколько лет при таком сценарии может быть потерян.
В остальном граждане не увидят разницы между хранением денег в ПФР или в НПФ. И в первом, и во втором случае деньги будут выплачиваться после достижения возраста 55 и 60 лет для женщин и мужчин соответственно. Даже условия наследования пенсии не отличаются. Если гражданину была назначена пожизненная выплата накопительной пенсии, его правопреемники не смогут ее получить. В остальных случаях пенсионные накопления наследуются по закону вне зависимости от того, хранились они в государственном или частном фонде.
В остальных случаях пенсионные накопления наследуются по закону вне зависимости от того, хранились они в государственном или частном фонде.
Выбирая негосударственный пенсионный фонд, обратите внимание на историю, репутацию НПФ и его показатели доходности. В частности, рекомендуем вам проверить:
- наличие лицензии. НПФ не может вести свою деятельность без лицензии, поэтому если у организации, которая представляется НПФ, ее нет – перед вами мошенники. Список лицензированных фондов размещен на сайте Банка России;
- присутствие НПФ в системе гарантирования прав застрахованных лиц. Все фонды, работающие в системе ОПС, обязаны быть участниками этой системы;
- актуальный рейтинг НПФ, который обновляется 2 раза за год. Это позволит оценить риски: наивысший рейтинг (ruAAA) с высокой вероятностью говорит о финансовой стабильности, низкий – о потенциальной неустойчивости. Имейте в виду, что не все НПФ дали согласие рейтинговому агентству на оценку – ряд фондов ее не имеют;
- величина собственных средств (капитала).
 Чем больше, тем лучше. Такой рэнкинг составляет «Национальное рейтинговое агентство»;
Чем больше, тем лучше. Такой рэнкинг составляет «Национальное рейтинговое агентство»; - количество застрахованных лиц и объем средств под управлением. Большое количество клиентов и их денег косвенно говорит о доверии к фонду (но, разумеется, успеха не гарантирует). Посмотреть ТОП-10 НПФ по объему средств под управлением вы можете в разделе «Доходность»;
- доходность за прошлые периоды. Об эффективности работы НПФ можно судить по тому, какой доход он приносит своим участникам. Причем смотреть доходность лучше не за один год, а за несколько. Если на протяжении ряда лет показатели доходности держатся выше инфляции, это хороший знак. Провести сравнение вы можете все в том же разделе «Доходность».
Можно ли вернуться в ПФР?Важно помнить, что ни один игрок- частный или государственный не может гарантировать доходность будущих периодов. И высокие результаты в прошлом не гарантируют успеха в будущем.
Если доходность в фонде вас все же не устраивает, можно выбрать новый НПФ или вернуться в ПФР. Причем неограниченное количество раз. Однако при переходе от одного страховщика к другому нужно помнить о «правиле пяти лет»: без потерь перевести деньги можно один раз в пятилетку. В противном случае есть риск остаться без инвестиционного дохода.
Причем неограниченное количество раз. Однако при переходе от одного страховщика к другому нужно помнить о «правиле пяти лет»: без потерь перевести деньги можно один раз в пятилетку. В противном случае есть риск остаться без инвестиционного дохода.
Подробнее – читайте статью «Как избежать потерь при смене страховщика?».
Статья 9. Основы взаимодействия Агентства, Банка России и Пенсионного фонда Российской Федерации / КонсультантПлюс
Статья 9. Основы взаимодействия Агентства, Банка России и Пенсионного фонда Российской Федерации
1. Агентство, Банк России и Пенсионный фонд Российской Федерации координируют свою деятельность и информируют друг друга по вопросам осуществления мероприятий по гарантированию прав застрахованных лиц, в том числе путем направления документов, в которых информация представлена в электронной форме и подписана усиленной квалифицированной электронной подписью.
(в ред. Федерального закона от 21.07.2014 N 218-ФЗ)
2. В целях прогнозирования и информационного обеспечения функционирования системы гарантирования прав застрахованных лиц Банк России направляет в Агентство отчетность фондов-участников, а также иные документы и информацию, касающиеся деятельности страховщиков, по запросу Агентства.
В целях прогнозирования и информационного обеспечения функционирования системы гарантирования прав застрахованных лиц Банк России направляет в Агентство отчетность фондов-участников, а также иные документы и информацию, касающиеся деятельности страховщиков, по запросу Агентства.
3. Банк России не позднее рабочего дня, следующего за днем принятия соответствующего решения, информирует Агентство и Пенсионный фонд Российской Федерации:
1) о вынесении положительного заключения о соответствии негосударственного пенсионного фонда требованиям статьи 19 настоящего Федерального закона;
(п. 1 в ред. Федерального закона от 29.06.2015 N 210-ФЗ)
2) о принятии решения о проведении проверки фонда-участника;
3) об аннулировании лицензии;
4) о переоформлении лицензии;
5) о введении запрета на проведение всех или части операций негосударственного пенсионного фонда (фонда-участника), а также запрета, предусмотренного статьями 21 и 22 настоящего Федерального закона;
6) о выдаче предписания об устранении фондом-участником выявленных нарушений требований законодательства Российской Федерации;
7) о реорганизации и ликвидации фонда-участника.
4. Агентство и Пенсионный фонд Российской Федерации вправе обращаться в Банк России с предложениями:
1) о проведении проверки фонда-участника Банком России. Порядок участия служащих Агентства в таких проверках, их права и обязанности устанавливаются нормативным актом Банка России по согласованию с Агентством;
2) о применении Банком России к фонду-участнику мер ответственности, предусмотренных федеральными законами, с приложением документов, обосновывающих необходимость применения указанных мер.
5. О принятом решении о проведении проверки фонда-участника Банком России и о применении Банком России к фонду-участнику мер ответственности, предусмотренных федеральными законами, Банк России сообщает Агентству и Пенсионному фонду Российской Федерации в течение пятнадцати календарных дней со дня получения соответствующего предложения, предусмотренного частью 4 настоящей статьи.
Открыть полный текст документа
Нормативные документы | Сайт отделения ПФР и Союза пенсионеров в Кировской области
 12.2001 N 167-ФЗ (ред. от 13.07.2015)
12.2001 N 167-ФЗ (ред. от 13.07.2015)«Об обязательном пенсионном страховании в Российской Федерации»
«О страховых взносах в Пенсионный фонд Российской Федерации, Фонд социального страхования Российской Федерации, Федеральный фонд обязательного медицинского страхования»
«Об организации предоставления государственных и муниципальных услуг»
«О персональных данных»
«Об электронной подписи»
«Об индивидуальном ( персонифицированном ) учете в системе обязательного пенсионного страхования «
«О дополнительных страховых взносах на накопительную часть трудовой пенсии и государственной поддержке формирования пенсионных накоплений «
«О перечне документов, представляемых в экспертный совет по технико-внедренческим особым экономическим зонам для получения заключения, подтверждающего, что организация оказывает инжиниринговые услуги в области высоких технологий»
 N 2п
N 2пОб утверждении формы расчета по начисленным и уплаченным страховым взносам на обязательное пенсионное страхование в пенсионный фонд российской федерации и на обязательное медицинское страхование в федеральный фонд обязательного медицинского страхования плательщиками страховых взносов, производящими выплаты и иные вознаграждения физическим лицам, и порядка ее заполнения
Об утверждении форм документов, используемых для регистрации граждан в системе обязательного пенсионного страхования, и Инструкции по их заполнению
«Об утверждении форм документов, применяемых при осуществлении контроля за уплатой страховых взносов»
«Об утверждении правил указания информации в реквизитах распоряжений о переводе денежных средств в уплату платежей в бюджетную систему Российской Федерации»
 12.2013 N 125н
12.2013 N 125н«Об утверждении порядка учета Федеральным казначейством поступлений в бюджетную систему Российской Федерации и их распределения между бюджетами бюджетной системы Российской Федерации»
«О порядке кассового обслуживания исполнения федерального бюджета, бюджетов субъектов Российской Федерации и местных бюджетов и порядке осуществления территориальными органами Федерального казначейства отдельных функций финансовых органов субъектов Российской Федерации и муниципальных образований по исполнению соответствующих бюджетов»
«О порядке сообщения банком в электронном виде органу контроля за уплатой страховых взносов об открытии или о закрытии счета, об изменении реквизитов счета»
«О порядке сообщения банком в электронном виде налоговому органу об открытии или о закрытии счета, об изменении реквизитов счета»
 08.2006 N 197п
08.2006 N 197п«Об утверждении порядка регистрации в территориальных органах ПФР страхователей, уплачивающих страховые взносы в ПФРФ в виде фиксированных платежей»
«Об утверждении порядка регистрации и снятия с рег.учета в территориальных органах ПФРФ страхователей, производящих выплаты физическим лицам»
«Об утверждении Правил подачи заявления о добровольном вступлении в правоотношения по обязательному пенсионному страхованию»
Об утверждении инструкции о порядке ведения индивидуального (персонифицированного) учета сведений о застрахованных лицах
«Об утверждении формы «Сведения о страховом стаже застрахованных лиц (СЗВ-СТАЖ)», формы «Сведения по страхователю, передаваемые в ПФР для ведения индивидуального (персонифицированного) учета (ОДВ-1)», формы «Данные о корректировке сведений, учтенных на индивидуальном лицевом счете застрахованного лица (СЗВ-КОРР)», формы «Сведения о заработке (вознаграждении), доходе, сумме выплат и иных вознаграждений, начисленных и уплаченных страховых взносах, о периодах трудовой и иной деятельности, засчитываемых в страховой стаж застрахованного лица (СЗВ-ИСХ)», порядка их заполнения и формата сведений»
 12.2015 № 511-п
12.2015 № 511-п«Об утверждении формы «Сведения о страховом стаже застрахованных лиц (СЗВ-СТАЖ)», формы «Сведения по страхователю, передаваемые в ПФР для ведения индивидуального (персонифицированного) учета (ОДВ-1)», формы «Данные о корректировке сведений, учтенных на индивидуальном лицевом счете застрахованного лица (СЗВ-КОРР)», формы «Сведения о заработке (вознаграждении), доходе, сумме выплат и иных вознаграждений, начисленных и уплаченных страховых взносах, о периодах трудовой и иной деятельности, засчитываемых в страховой стаж застрахованного лица (СЗВ-ИСХ)», порядка их заполнения и формата сведений»
«О внедрении защищенного электронного документооборота в целях реализации законодательства российской федерации об обязательном пенсионном страховании»
«Об утверждении методических рекомендаций по организации проведения выездных проверок плательщиков страховых взносов»
 N 35п
N 35п«Об утверждении форм документов,применяемых при предоставлении отсрочки(рассрочки)по уплате страховых взносов, пеней и штрафов»
О внесении изменений в постановление правления пенсионного фонда Российской Федерации от 16 января 2014 года N 2П
О предельной величине базы для начисления страховых взносов в Фонд социального страхования Российской Федерации и Пенсионный фонд Российской Федерации с 1 января 2015г.
«Об утверждении формата данных сведений о застрахованных лицах»
«Об утверждении Административного регламента предоставления Пенсионным фондом Российской Федерации государственной услуги по информированию зарегистрированных лиц о состоянии их индивидуальных лицевых счетов в системе обязательного пенсионного страхования согласно Федеральным законам «Об индивидуальном (персонифицированном) учете в системе обязательного пенсионного страхования» и «Об инвестировании средств для финансирования накопительной пенсии в Российской Федерации»
 02.2019 № 38п
02.2019 № 38п«Об утверждении Административного регламента предоставления Пенсионным фондом Российской Федерации государственной услуги по приему от застрахованных лиц заявлений о добровольном вступлении в правоотношения по обязательному пенсионному страхованию в целях уплаты дополнительных страховых взносов на накопительную пенсию в соответствии с Федеральным законом от 30 апреля 2008 г. N 56-ФЗ «О дополнительных страховых взносах на накопительную пенсию и государственной поддержке формирования пенсионных накоплений»
«Об утверждении Административного регламента предоставления Пенсионным фондом Российской Федерации государственной услуги по приему от граждан анкет в целях регистрации в системе индивидуального (персонифицированного) учета, в том числе по приему от зарегистрированных лиц заявлений об изменении анкетных данных, содержащихся в индивидуальном лицевом счете, или о выдаче документа, подтверждающего регистрацию в системе индивидуального (персонифицированного) учета»
 05.2019 N 277п
05.2019 N 277п«Об утверждении Административного регламента предоставления Пенсионным фондом Российской Федерации государственной услуги по приему от плательщиков страховых взносов расчетов по начисленным и уплаченным страховым взносам на обязательное пенсионное страхование и обязательное медицинское страхование»
«Об утверждении формы расчета по начисленным и уплаченным страховым взносам на обязательное пенсионное страхование в Пенсионный фонд Российской Федерации и страховым взносам на обязательное медицинское страхование в Федеральный фонд обязательного медицинского страхования главами крестьянских (фермерских) хозяйств и Порядка ее заполнения»
«Об утверждении Административного регламента предоставления Пенсионным фондом Российской Федерации государственной услуги по приему от страхователей реестров застрахованных лиц о перечислении дополнительных страховых взносов на накопительную пенсию в соответствии с Федеральным законом «О дополнительных страховых взносах на накопительную пенсию и государственной поддержке формирования пенсионных накоплений»
 01.2020 № 46па
01.2020 № 46па«Об утверждении формы сведений о трудовой деятельности, представляемой работнику работодателем, формы предоставления сведений о трудовой деятельности из информационных ресурсов Пенсионного фонда Российской Федерации и порядка их заполнения»
 08.2020 № 533н
08.2020 № 533н«Об утверждении особенностей представления сведений о трудовой деятельности государственными органами в отношении отдельных категорий зарегистрированных лиц;
«О внесении изменений в Административный регламент предоставления Пенсионным фондом Российской Федерации государственной услуги по приему от плательщиков страховых взносов расчетов по начисленным и уплаченным страховым взносам на обязательное пенсионное страхование и обязательное медицинское страхование, утвержденный постановлением Правления ПФР от 17 мая 2019 гола № 277п»
Взаимодействие НПФ и ПФР в СМЭВ: сверка квитанций
В материале содержится программа для сопоставления исходящих СМЭВ-запросов негосударственного пенсионного фонда и квитанций, полученных в ответ от Пенсионного фонда России. А также приводится инструкция и примеры использования утилиты.
Отчеты НПФ и квитанции ПФР в СМЭВ
Негосударственные пенсионные фонды через СМЭВ интенсивно обмениваются большим количеством разнообразных документов и отчетов с Пенсионным фондом России. Практически на каждый документ, отправленный НПФом, ПФР должен подготовить встречный документ с результатами проверки и приемки отчетного документа на стороне НПФ (назовем такой документ квитанцией).
Практически на каждый документ, отправленный НПФом, ПФР должен подготовить встречный документ с результатами проверки и приемки отчетного документа на стороне НПФ (назовем такой документ квитанцией).
Обмен отчетными документами и квитанциями осуществляется посредством передачи файлов-вложений в рамках видов сведений «Входящие документы из НПФ в ПФР» и «Исходящие документы из ПФР в НПФ». При этом участникам межведомственного взаимодействия необходимо сопоставлять отчеты, направленные через один вид сведений, с квитанциями, полученными посредством второго ВС.
Стандартные возможности СМЭВ-адаптера позволяют сопоставлять сообщения и вложения, но не позволяют идентифицировать, квитанцией к какому документу является полученное вложение.
Сопоставление СМЭВ-отчетов и квитанций
Для того чтобы облегчить поиск и сопоставление ответов на запросы в ПФР команда Хемуль IT разработала для своих клиентов простую утилиту на Java. Утилита обходит все каталоги с файлами вложений, распаковывает их и извлекает из каждого файла его метаданные (элементы «GUI», «ВОтветНа» и «ДатаВремя» из блока «СлужебнаяИнформация»).
Метаданные вместе с названием каталога и именем файла сохраняются в таблице (CSV-файле), которая затем обрабатывается вручную средствами MS Excel.
Порядок установки и использования ПО
Предусловие: наличие JRE версии от 1.8 и выше.
Прилагаемый архив необходимо распаковать в отдельный каталог, например: /opt/report (или C:\report).
В файле config.ini нужно задать два параметра:
DOC_DIR= /opt/smev/date/base-storage/XXYYZZ
(или C:\\SmevAdapter\\data\\base-storage\\XXYYZZ) — корневой каталог, в который СМЭВ-адаптер помещает файлы вложенийREPORT_FILE=./npf_report.csv(или C:\\Temp\\npf_report.csv) — путь и имя csv-файла с отчетом.
Утилита запускается в консоли командой:
java -jar npf-report-1.1.jar
В процессе работы могут появляться сообщения о файлах нулевой длины, битых архивах, которые невозможно распаковать, и некорректных XML-файлах, которые невозможно распарсить. Такие файлы пропускаются и не влияют на обработку оставшихся файлов.
Такие файлы пропускаются и не влияют на обработку оставшихся файлов.
По завершении работы в консоль будет выведена надпись «Обработка всех вложений завершена, отчет сформирован».
Работа с отчетом
Результатом работы утилиты является файл в формате *.csv, который может быть обработан далее вручную по следующему сценарию:
- Фиксируем первую строку с заголовками и добавляем фильтр.
- В фильтре на столбце «ВОтветНа» выбираем значение «parsing error» и получаем список «кривых» файлов, которые не удалось обработать.
При желании можно скопировать этот список в отдельную таблицу, чтобы потом разобраться с причинами ошибок.
Но в итоге эти строки нужно удалить из нашего отчета. - Теперь в фильтре на столбце «ВОтветНа» выбираем флажок «Выделить все» и снимаем флажок «(Пустые)».
Получаем список всех файлов, в которых присутствует элемент «ВОтветНа», т.е. список квитанций.
Создаем новый лист в этой же книге Excel и копируем туда список квитанций.
- Удаляем строки квитанций с первого листа Excel.
- Удаляем столбец «ВОтветНа».
- Столбцы E, F, G, H называем соответственно «Каталог квитанции», «Файл квитанции», «GUID квитанции», «ДатаВремя квитанции».
- В ячейке E2 вводим формулу «=ВПР($C2;Лист1!$A:$E;2;ЛОЖЬ)»
В ячейке F2 вводим формулу «=ВПР($C2;Лист1!$A:$E;3;ЛОЖЬ)»
В ячейке G2 вводим формулу «=ВПР($C2;Лист1!$A:$E;4;ЛОЖЬ)»
В ячейке h3 вводим формулу «=ВПР($C2;Лист1!$A:$E;5;ЛОЖЬ)» - Ячейки E2:h3 копируем вниз до конца заполненного диапазона.
- В фильтре на столбце E выбираем флажок «Выделить все» и снимаем флажок «#Н/Д».
Получаем список всех файлов, на которые от ПФР пришли квитанции.
Если инвертировать условия фильтрации, то получим список файлов, на которые от ПФР не пришли квитанции. - Сохраняем получившийся отчет в формате xls.
По ссылке можно посмотреть пример преобразованного файла.
0 0 голоса
Рейтинг статьи
Актуальная информация от Пенсионного фонда РФ
Перевести накопительную пенсию из ПФР в НПФ и обратно или поменять один НПФ на другой можно в клиентских службах Пенсионного фонда или через портал Госуслуг
Приём заявлений граждан о переходе в негосударственный пенсионный фонд (НПФ) или Пенсионный фонд России (ПФР), включая досрочный переход, а также уведомлений о замене страховщика и отказе от смены страховщика осуществляется двумя способами:
— в форме электронного документа через Портал госуслуг;
— путём личного обращения (через представителя) в клиентские службы ПФР.
Напомним, что у граждан, имеющих пенсионные накопления, есть право доверить их управление:
— Пенсионному фонду России, выбрав управляющую компанию (УК), отобранную по кон-курсу, с которой ПФР заключил договор доверительного управления средствами пенсионных накоплений, в том числе один из инвестиционных портфелей государственной управляющей компании (ГУК) — ВЭБ.РФ;
— негосударственному пенсионному фонду, осуществляющему деятельность по обязательно-му пенсионному страхованию.
Менять страховщика (ПФР или НПФ), управляющую компанию можно ежегодно. Но при этом важно помнить, что если гражданин будет осуществлять смену страховщика чаще одного раза в пять лет, он может потерять инвестиционный доход, полученный предыдущим страховщиком. Если же страховщиком гражданина является ПФР, смену управляющей ком-пании можно производить ежегодно без потери инвестиционного дохода.
В случае, если гражданин выбирает вариант досрочного перехода к новому страхов-щику, при подаче заявления он в обязательном порядке информируется Пенсионным фондом о сумме инвестиционного дохода, которую он при этом потеряет.
Данная информация позволит гражданину взвесить все «за» и «против» и сделать осо-знанный выбор, согласен ли он потерять инвестиционный доход при досрочной смене стра-ховщика или стоит подать заявление со сроком перехода через 5 лет. Вне зависимости от вида заявления ПФР будет сообщать текущему страховщику и новому страховщику, кото-рый указан в заявлении гражданина, о факте подачи им заявления или уведомления.
Обращаем внимание, что если вы планируете перейти в НПФ, то до подачи заявления необходимо заключить договор с выбранной организацией. Сделать это можно не позднее 1 декабря текущего года.
С перечнем негосударственных пенсионных фондов и управляющих компаний можно ознакомиться на официальном сайте ПФР www.pfrf.ru.
___________________________________________________________________________________________________________________
Идентификация членов семейства факторов, взаимодействующих с фитохромами, и функциональный анализ MdPIF4 у Malus domestica
Abstract
Факторы, взаимодействующие с фитохромами (PIF), члены семейства основных транскрипционных факторов спираль-петля-спираль, которые были тщательно исследованы у Arabidopsis thaliana , играют важную роль в росте и развитии растений. Тем не менее, членов PIF не были систематически исследованы на яблоках, многолетней древесной культуре, имеющей экономическое значение во всем мире.Здесь семь генов PIF были идентифицированы из эталонного генома Malus × domestica . Были проанализированы хромосомные местоположения, структуры генов и филогенетические отношения этих членов. Анализ цис-действующих элементов в промоторных областях генов MdPIF показал, что различные элементы связаны со светом, абиотическим стрессом и чувствительностью растений к гормонам. Впоследствии анализ субклеточной локализации и транскрипционной активности показал, что MdPIFs являются типичными ядерными факторами транскрипции со способностью активации транскрипции.Анализ экспрессии показал, что генов MdPIF имели разные паттерны экспрессии генов для различных абиотических факторов. Более того, сверхэкспрессированный MdPIF4 снижал чувствительность каллусов яблони к абсцизовой кислоте (АБК).
Тем не менее, членов PIF не были систематически исследованы на яблоках, многолетней древесной культуре, имеющей экономическое значение во всем мире.Здесь семь генов PIF были идентифицированы из эталонного генома Malus × domestica . Были проанализированы хромосомные местоположения, структуры генов и филогенетические отношения этих членов. Анализ цис-действующих элементов в промоторных областях генов MdPIF показал, что различные элементы связаны со светом, абиотическим стрессом и чувствительностью растений к гормонам. Впоследствии анализ субклеточной локализации и транскрипционной активности показал, что MdPIFs являются типичными ядерными факторами транскрипции со способностью активации транскрипции.Анализ экспрессии показал, что генов MdPIF имели разные паттерны экспрессии генов для различных абиотических факторов. Более того, сверхэкспрессированный MdPIF4 снижал чувствительность каллусов яблони к абсцизовой кислоте (АБК).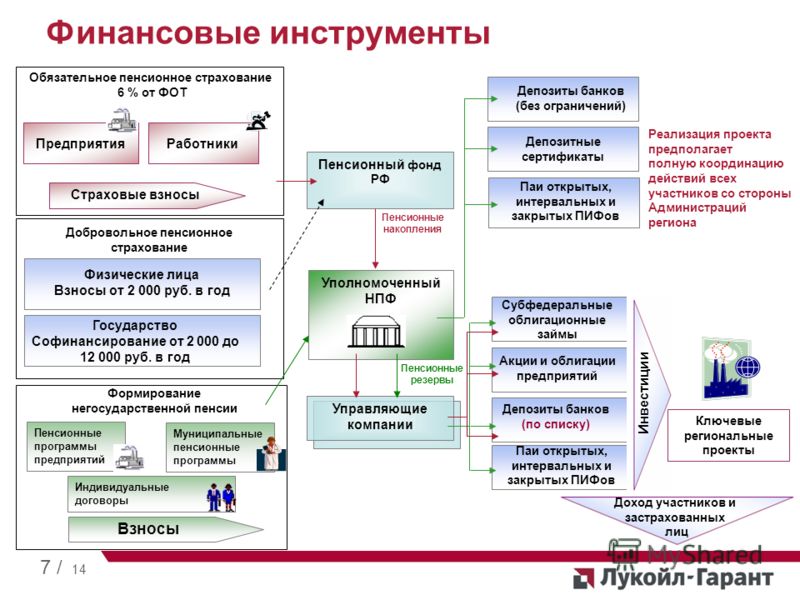 Наша работа закладывает основу для дальнейшего изучения функций PIF в росте и развитии растений яблони.
Наша работа закладывает основу для дальнейшего изучения функций PIF в росте и развитии растений яблони.
Ключевые слова: фитохром-взаимодействующий фактор, анализ экспрессии генов, локализация белка, чувствительность к АБК, активность активации транскрипции
1.Введение
Свет является одним из важнейших факторов внешней среды, влияющих на рост и развитие растений [1]. Из-за различий в спектре света, интенсивности, направлении и фотопериоде ряд фоторецепторов, включая фитохром, криптохром и фототропин, эволюционировал для адаптации растений к различным условиям освещения [1,2,3]. Фитохром представляет собой важный класс фоторецепторов, которые воспринимают и реагируют на красный и дальний красный свет и участвуют во всем процессе роста и развития, от прорастания семян до вегетативного роста и созревания растений [4,5].На сегодняшний день у Arabidopsis идентифицировано пять фитохромов (phyA-phyE), и они имеют две фото-взаимопревращаемые формы [6,7]. При воздействии красного света конформация фитохромов аллостерически изменяется от неактивной формы, поглощающей красный свет (Pr), до активной формы, поглощающей дальний красный свет (Pfr). Неактивная форма Pr расположена в цитозоле, тогда как активная форма Pfr находится в ядре [6,7]. При перемещении в ядро фитохромы в активной форме взаимодействуют с множеством факторов, регулируя транскрипцию нижестоящих генов-мишеней и опосредуя последующие управляемые фотореакции [8,9,10,11].
При воздействии красного света конформация фитохромов аллостерически изменяется от неактивной формы, поглощающей красный свет (Pr), до активной формы, поглощающей дальний красный свет (Pfr). Неактивная форма Pr расположена в цитозоле, тогда как активная форма Pfr находится в ядре [6,7]. При перемещении в ядро фитохромы в активной форме взаимодействуют с множеством факторов, регулируя транскрипцию нижестоящих генов-мишеней и опосредуя последующие управляемые фотореакции [8,9,10,11].
Факторы, взаимодействующие с фитохромами (PIFs), являются одним из наиболее важных партнеров, взаимодействующих с фитохромами, и они в первую очередь действуют как негативные регуляторы фотоморфогенеза в ответ на свет и поддерживают скотоморфогенез в темноте [8,12]. В настоящее время восемь PIF (AtPIF1–AtPIF8) идентифицированы у Arabidopsis [9], а AtPIF3 первоначально был идентифицирован как белок, взаимодействующий с фитохромом, с использованием AtphyB в качестве приманки [13]. Являясь членами семейства основных транскрипционных факторов спираль-петля-спираль (bHLH), все AtPIF содержат домен bHLH, который играет ключевую роль в образовании гомо- и гетеродимеров AtPIF [9]. Кроме того, мотив активного связывания phyB (APB) присутствует во всех AtPIF, тогда как мотив активного связывания phyA (APA) существует только в AtPIF1 и AtPIF3 [8,14,15].
Кроме того, мотив активного связывания phyB (APB) присутствует во всех AtPIF, тогда как мотив активного связывания phyA (APA) существует только в AtPIF1 и AtPIF3 [8,14,15].
В дополнение к тому, что PIF являются негативным регулятором фотоморфогенеза в ответ на свет [8,16,17], также было показано, что PIF регулируют многие другие пути, включая биосинтез антоцианов, термоморфогенез, передачу сигналов гормонов и реакцию на биотические и абиотические стрессы. путем взаимодействия с несколькими клеточными молекулами [18,19,20,21].Например, кукуруза ( Zea mays ) ZmPIF1 и ZmPIF3 могут улучшить засухоустойчивость риса, индуцируя закрытие устьиц [22,23]. AtPIF1 взаимодействует с AtABI5 (нечувствительным к абсцизовой кислоте 5), фактором транскрипции основной лейциновой молнии (bZIP), который участвует в сигнальном пути абсцизовой кислоты (ABA), для регулирования эндогенных сигнальных путей ABA и ABA [24,25]. Кроме того, рост гипокотилей мутанта pif4 нечувствителен к высокой температуре (29 °C), что указывает на ключевую роль AtPIF4 в термореактивном росте растений [26]. Кроме того, сообщалось, что AtPIF4 ускоряет цветение путем прямой активации экспрессии AtFT ( flowing locus T ) в зависимости от температуры [27]. Таким образом, PIF считаются центром интеграции экологических и гормональных сигнальных путей [28].
Кроме того, сообщалось, что AtPIF4 ускоряет цветение путем прямой активации экспрессии AtFT ( flowing locus T ) в зависимости от температуры [27]. Таким образом, PIF считаются центром интеграции экологических и гормональных сигнальных путей [28].
Яблоня ( Malus domestica ) является важной товарной плодовой культурой, и на ее производство сильно влияют факторы окружающей среды, особенно свет [29,30]. PIF играют центральную роль в адаптации растений к различным условиям освещения; однако идентификация и функциональный анализ PIF в яблоках систематически не исследовались, за исключением MdPIF1 [31].Здесь мы идентифицировали семь генов MdPIF с использованием методов биоинформатики и проанализировали их генные структуры, расположение в хромосомах, консервативные мотивы и цис-действующие элементы их промоторных областей. Паттерны экспрессии при различных стрессах были проанализированы, чтобы лучше понять функцию генов Apple PIF . Более того, сверхэкспрессированный MdPIF4 снижал чувствительность каллусов яблони к АБК. Наши результаты создают основу для дальнейшего функционального исследования PIF растений.
Более того, сверхэкспрессированный MdPIF4 снижал чувствительность каллусов яблони к АБК. Наши результаты создают основу для дальнейшего функционального исследования PIF растений.
2. Результаты
2.1. Идентификация, хромосомная локализация и анализ структуры генов MdPIF Гены
PIF являются членами подмножества надсемейства факторов транскрипции bHLH. Сообщалось, что яблоки имеют 188 генов MdbHLH [32]. Чтобы идентифицировать членов PIF в яблоках, мы использовали белковых последовательностей PIF Arabidopsis для проведения анализа BLASTp, и восемь кандидатов MdPIF (названные от MdPIF1 до MdPIF8) были получены из яблок.В зависимости от наличия консервативных доменов bHLH и APB окончательно определили семь белков MdPIF, а MdPIF6 (инвентарный номер в GenBank: MDP0000215587) исключили из-за отсутствия домена APB (и рис. S1). Анализ последовательности показал, что эти белки MdPIF широко различаются по длине, от 449 аминокислот (аа) до 1040 аа, а их молекулярная масса колеблется от 48 кДа до 113 кДа. Другие характеристики, включая идентификаторы генов, геномные положения, общее среднее гидропатичности и значения pI белков MdPIF, суммированы в .
Другие характеристики, включая идентификаторы генов, геномные положения, общее среднее гидропатичности и значения pI белков MdPIF, суммированы в .
Таблица 1
Информация о членах PIF , обнаруженных в яблоках.
| Gene Name | Gene | IDРасположение Chromosome | позиция | MRNA Длина (BP) | CDS Длина (BP) | Длина аминокислота (AA) | молекулярному вес Da) | Теоретическая pI | Общее среднее гидропатичности (GRAVY) | Лучшие совпадений | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| МдПИФ1 | MDP0000289642 | chr10 | 16985872-16992614 | 6743 | 1920 | 639 | 628,55 | −0,59 | АтПИФ1 | ||
| МдПИФ2 | MDP0000205358 | CHR4 | 18339668-18342979 | 3312 | 2935 | 744 | 80495. 75 75 | 6.52 | -0.645 | АтПИФ2 | |
| МдПИФ3 | MDP00002 | chr12 | 27035201-27038407 | 3207 | 2127 | 708 | 445,83 | −0,626 | АтПИФ3 | ||
| МдПИФ4 | MDP0000198404 | CHR17 | 11357450-11361687 | 4238 | 1842 | 613 | 67336. 57 57 | 67336.57 | 7.06 | —0.676 | АтПИФ4 |
| МдПИФ5 | MDP0000254650 | chr9 | 11107477-11118877 | 11401 | 3123 | 1040 | 129 | 8,76 | −0,52 | АтПИФ5 | |
| МдПИФ7 | MDP0000319248 | CHR14 | 26344135-26351096 | 6962 | 1746 | 1746 | 581 | 64713. 13 13 | 9.96 | —0.903 | АтПИФ7 |
| МдПИФ8 | MDP0000439540 | chr7 | 10087113-10092308 | 5196 | 1350 | 449 | 517,72 | −0,445 | АтПИФ8 |
На основе информации о местоположении генома, полученной из базы данных геномов розоцветных (ГДР), генов MdPIF были картированы на семи хромосомах яблони (Chr): Chr 4, Chr 7, Chr 9, Chr 10, Chr 12, Хр 14 и Хр 17 соответственно (а). Анализируя последовательности геномной ДНК, мы обнаружили, что количество интронов этих MdPIF варьировалось от 5 до 18. MdPIF5 содержал наибольшее количество интронов (18), тогда как MdPIF8 имел только пять интронов.Одна группа из генов MdPIF ( MdPIF4 и 7 ) содержала одинаковое количество интронов/экзонов (б). Кроме того, MdPIF2 и MdPIF3 , оба из которых были расположены в одной и той же ветви филогенетического дерева, построенного на основе кодирующих последовательностей MdPIF (рис. S2), имели сходное распределение интронов/экзонов.
Анализируя последовательности геномной ДНК, мы обнаружили, что количество интронов этих MdPIF варьировалось от 5 до 18. MdPIF5 содержал наибольшее количество интронов (18), тогда как MdPIF8 имел только пять интронов.Одна группа из генов MdPIF ( MdPIF4 и 7 ) содержала одинаковое количество интронов/экзонов (б). Кроме того, MdPIF2 и MdPIF3 , оба из которых были расположены в одной и той же ветви филогенетического дерева, построенного на основе кодирующих последовательностей MdPIF (рис. S2), имели сходное распределение интронов/экзонов.
Хромосомное расположение и структура генов (интрон/экзон) генов MdPIF . ( a ) Хромосомное расположение семи генов MdPIF на семи хромосомах яблока.( b ) Генные структуры генов MdPIF . Красные прямоугольники обозначают экзоны, а синие линии — интроны.
2.2. Филогенетический анализ, множественное выравнивание последовательностей и предсказание консервативных мотивов MdPIF
Поскольку PIF являются факторами транскрипции bHLH, которые сохраняются среди различных видов растений, мы затем провели филогенетический анализ, основанный на белковых последовательностях PIF из яблока, Arabidopsis и рис ( Oryza sativa ), чтобы исследовать их эволюционные отношения. Мы обнаружили, что белки MdPIF были разделены на четыре группы. MdPIF1 принадлежал к группе I; MdPIF4 и MdPIF5 были во II группе; MdPIF2 и MdPIF3 были отнесены к группе III; а MdPIF7 и MdPIF8 были отнесены к группе IV (а). Более того, MdPIF имели более тесную связь с AtPIF, чем с OsPIF, что, вероятно, было связано с тем, что и яблоня, и видов Arabidopsis являются двудольными, тогда как виды риса являются однодольными.
Мы обнаружили, что белки MdPIF были разделены на четыре группы. MdPIF1 принадлежал к группе I; MdPIF4 и MdPIF5 были во II группе; MdPIF2 и MdPIF3 были отнесены к группе III; а MdPIF7 и MdPIF8 были отнесены к группе IV (а). Более того, MdPIF имели более тесную связь с AtPIF, чем с OsPIF, что, вероятно, было связано с тем, что и яблоня, и видов Arabidopsis являются двудольными, тогда как виды риса являются однодольными.
Анализ филогенетического родства и консервативных доменов MdPIF.( a ) Неукорененное филогенетическое дерево белков PIF из Malus domestica , Arabidopsis thaliana и Oryza sativa. MEGA5.1 использовали для построения филогенетического дерева на основе последовательностей белков PIF, и показаны четыре отдельные подгруппы белков PIF. Онлайн-программное обеспечение Interactive Tree of Life (iTOL) использовалось для аннотирования и обзора филогенетического дерева. AtPIF1: AT2G20180; AtPIF2: AT2G46970; AtPIF3: AT1G09530; AtPIF4: AT2G43010; AtPIF5: AT3G59060; AtPIF6: AT3G62090; AtPIF7: AT5G61270; AtPIF8: AT4G00050; ОсПИЛ11: Ос12г0610200; ОсПИЛ12: Ос03г0639300; ОсПИЛ13: Ос03г0782500; ОсПИЛ14: Ос07г0143200; ОсПИЛ15: Ос01г0286100; ОсПИЛ16: Ос05г0139100. ( b ) Сравнение мотивов белков MdPIF и AtPIF. Наличие мотивов APA, APB и bHLH показано прямоугольниками. Бар = 100 аминокислотных остатков.
( b ) Сравнение мотивов белков MdPIF и AtPIF. Наличие мотивов APA, APB и bHLH показано прямоугольниками. Бар = 100 аминокислотных остатков.
Наличие доменов bHLH и APB было разработано в качестве основного критерия для скрининга членов PIF. Здесь два высококонсервативных домена были идентифицированы во всех белках MdPIF (b и рисунок S3a). Кроме того, было обнаружено, что домен APA присутствует на N-конце нескольких белков MdPIF (MdPIF1, MdPIF2, MdPIF3, MdPIF4 и MdPIF5) (b и рисунок S3b), что сходно с AtPIF1 и AtPIF3 [8,15]. ].
Используя программу анализа мотивов MEME, мы предсказали 15 мотивов в белках MdPIF (4). Среди них мотив 1 (мотив HLH) и мотив 3 (мотив APB) широко распространены во всех белках MdPIF. Интересно, что мы обнаружили, что члены MdPIF в пределах одной и той же эволюционной ветви имеют сходное распределение мотивов. Например, MdPIF2 и MdPIF3, оба из которых расположены на одной и той же ветви, имеют очень сходный состав мотивов, и это также наблюдалось для MdPIF4 и MdPIF5 (10).
Схематическое изображение консервативных мотивов, предсказанных в белках MdPIF.Программа MEME использовалась для предсказания консервативных мотивов, а TBtools использовалась для демонстрации результатов.
2.3. Анализ цис-действующих элементов в промоутерах гена MdPIF
Сообщается, что PIF участвуют в реакциях растений на многочисленные абиотические факторы, и цис-действующие элементы в промоторной области играют решающую роль в этих процессах [8,33,34] . Таким образом, затем мы проанализировали цис-действующие элементы в промоторных областях генов MdPIF , чтобы изучить их потенциальные регуляторные паттерны (рис. S4).Результаты показали, что более 26 элементов распределены с промоторами генов MdPIF (таблица S2). В зависимости от их функции цис-действующие элементы были разделены на три группы: свето-, гормоно- и стресс-реактивные. Среди них светочувствительные элементы присутствовали во всех последовательностях промотора гена MdPIF , включая G-box (TACGAT) [35], ACE (CTAACGTATT), LAMP (CTTTATCA) и т.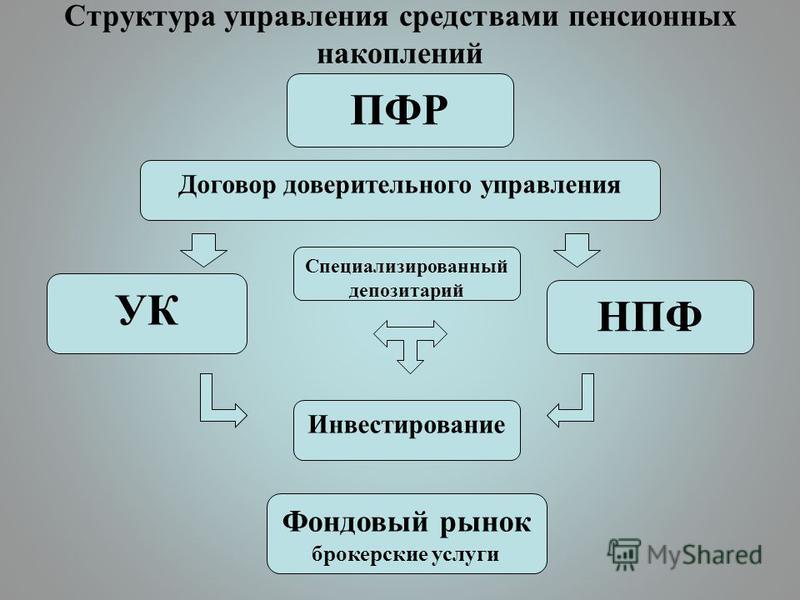 д. (таблица S2). Растительные гормональные элементы, такие как ABRE (ACGTG), GARE (TCTGTTG) и CGTCA, которые были вовлечены в реакцию на АБК, гиббереллин (GA) и жасмоновую кислоту (JA), соответственно, также были идентифицированы в промоторах генов MdPIF (таблица S2).Кроме того, было обнаружено множество элементов, реагирующих на абиотические стрессы (и Таблица S2). Например, промотор MdPIF7 содержал цис-действующие элементы, реагирующие на низкую температуру (LTR: CCGAAA) и раны (WUN-мотив: AAATTTCCT). В совокупности присутствие этих цис-действующих элементов указывает на то, что MdPIF могут быть вовлечены во множественные реакции, включая реакции на свет, гормоны и абиотические стрессы, для выяснения которых требуются дальнейшие усилия.
д. (таблица S2). Растительные гормональные элементы, такие как ABRE (ACGTG), GARE (TCTGTTG) и CGTCA, которые были вовлечены в реакцию на АБК, гиббереллин (GA) и жасмоновую кислоту (JA), соответственно, также были идентифицированы в промоторах генов MdPIF (таблица S2).Кроме того, было обнаружено множество элементов, реагирующих на абиотические стрессы (и Таблица S2). Например, промотор MdPIF7 содержал цис-действующие элементы, реагирующие на низкую температуру (LTR: CCGAAA) и раны (WUN-мотив: AAATTTCCT). В совокупности присутствие этих цис-действующих элементов указывает на то, что MdPIF могут быть вовлечены во множественные реакции, включая реакции на свет, гормоны и абиотические стрессы, для выяснения которых требуются дальнейшие усилия.
Таблица 2
Анализ цис-действующих элементов в промоторах гена MdPIF .
| Ген | Световая реакция | Гормон Ответ | Реакция на стресс | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ABA | ABA | GA | JA | SA | DT | LT | DNS | WD | AAI | ||
| МдПИФ1 | 4/1 | 1/1 | 1/0 | 0/1 | |||||||
| МдПИФ2 | 8/5 | 1/2 | 1/1 | 0/1 | 1/0 | 1/0 | |||||
| МдПИФ3 | 7/4 | 0/2 | 1/0 | 1/1 | 0/1 | 0/1 | 2/1 | ||||
| МдПИФ4 | 8/5 | 1/0 | 3/2 | 1/0 | 1/1 | 0/1 | |||||
| МдПИФ5 | 6/6 | 1/1 | 2/0 | 1/0 | 1/0 | 1/3 | |||||
| МдПИФ7 | 6/3 | 1/1 | 1/2 | 0/1 | 1/1 | 2/1 | |||||
| МдПИФ8 | 7/2 | 0/1 | 1/0 | 1/1 |
2.
 4. Профили экспрессии генов MdPIF
4. Профили экспрессии генов MdPIFPIF являются важными светочувствительными генами, поэтому далее мы проанализировали профили экспрессии всех генов MdPIF в ответ на суточный ритм. Сначала мы использовали qRT-PCR для определения уровней экспрессии генов MdPIF в различных органах, включая корень, стебель, лист, цветок и плод. Все генов MdPIF имели относительно сходные паттерны экспрессии. Все они имели самый высокий уровень экспрессии в листе и самый низкий уровень экспрессии в цветке и плоде (a-g), за исключением MdPIF7. имел самый низкий уровень экспрессии в стебле (f).
Профили экспрессии генов MdPIF . Профиль экспрессии MDPIF1 ( A ), MDPIF2 ( B ), MDPIF3 ( C ), MDPIF4 ( D ), MDPIF5 ( E ), MDPIF7 ( f ) и MdPIF8 ( г ) в различных тканях яблока. 18S рибосомную РНК использовали в качестве внутреннего контроля для анализа qRT-PCR. Данные представляют собой среднее значение ± стандартное отклонение для трех независимых биологических повторов.Звездочками отмечены достоверные различия по сравнению с тканью листа (*, p < 0,05; **, p < 0,01). ( h ) Профиль экспрессии MdPIF в зависимости от дня и ночи. Условия освещения от 8 до 24 часов, затем 8 часов темноты. Данные представляют собой среднее значение ± стандартное отклонение для трех независимых биологических повторов. Звездочками отмечены значимые отличия от контроля (*, p < 0,05; **, p < 0,01).
MdPIF1, 3, 4, 5, и 7 имели схожие паттерны экспрессии в ответ на суточный ритм.Уровни транскриптов пяти генов значительно увеличились за ночь и резко упали в течение дня. Среди них MdPIF5 имели пиковое изменение в 20:00. а его наивысший уровень был подобен тому, который наблюдался на рассвете. Однако уровень экспрессии MdPIF2 и MdPIF8 достиг пика в течение дня. MdPIF2 достигал максимального уровня в 12:00, затем быстро снижался и постепенно накапливался ночью. Экспрессия MdPIF8 снижалась с 16:00 ч.м. до 20:00 и повторно накапливается в течение ночи (h). В целом характер экспрессии генов MdPIF в ответ на суточные циклы был сходен с таковым у PIF4 и PIF5 как у Arabidopsis , так и у кукурузы, уровни транскриптов которых повышались ночью, достигали максимума на рассвете и упал в течение дня [36,37].
Однако уровень экспрессии MdPIF2 и MdPIF8 достиг пика в течение дня. MdPIF2 достигал максимального уровня в 12:00, затем быстро снижался и постепенно накапливался ночью. Экспрессия MdPIF8 снижалась с 16:00 ч.м. до 20:00 и повторно накапливается в течение ночи (h). В целом характер экспрессии генов MdPIF в ответ на суточные циклы был сходен с таковым у PIF4 и PIF5 как у Arabidopsis , так и у кукурузы, уровни транскриптов которых повышались ночью, достигали максимума на рассвете и упал в течение дня [36,37].
2.5. Субклеточная локализация и анализ транскрипционной активности MdPIF
Ядерная локализация является одной из ключевых общих черт транскрипционных факторов.Поэтому затем мы обнаружили субклеточную локализацию MdPIF1, MdPIF3, MdPIF4 и MdPIF8, которые были выбраны в качестве репрезентативных членов четырех групп на филогенетическом дереве (а). Конструкции MdPIF-зеленый флуоресцентный белок (GFP) вводили в листьев Nicotiana benthamiana посредством агроинфильтрации. После двухдневной обработки в темноте образцы исследовали под конфокальным микроскопом. Все MdPIF были исследованы в ядре (а), что согласуется с предыдущими сообщениями о том, что все PIF у арабидопсиса , и кукурузы были локализованы в ядре [12,38].
После двухдневной обработки в темноте образцы исследовали под конфокальным микроскопом. Все MdPIF были исследованы в ядре (а), что согласуется с предыдущими сообщениями о том, что все PIF у арабидопсиса , и кукурузы были локализованы в ядре [12,38].
Анализ субклеточной локализации и трансактивации белков MdPIF. ( a ) Субклеточная локализация белков MdPIF. Масштабная линейка = 20 мкм. ( b ) Схематическая диаграмма структуры MdPIF-pGBKT7. ( c ) Конструкцию pGBKT7-MdPIF трансформировали в дрожжевой штамм Ah209 и исследовали на чашках SD/-Trp и SD/-Trp/-His/-Ade/X-α-gal.
Транскрипционная активность белков MdPIF также была проверена на дрожжах. Кодирующие последовательности генов MdPIF были встроены в вектор pGBKT7 (b).Пустой вектор pGBKT7 и pGBKT7-MdMYB23 служили в качестве отрицательного и положительного контроля соответственно. Все дрожжи в составе разных конструкций нормально росли на среде SD/-Trp (в). После переноса в среду SD/-Trp/-His/-Ade с добавлением X-α-gal синие колонии дрожжей присутствовали при трансформации в pGBKT7-MdPIF или pGBKT7-MdMYB23, тогда как отрицательный контроль ничего не показывал. Эти результаты показали, что белки MdPIF характеризуются транскрипционной активностью в дрожжевых клетках.
После переноса в среду SD/-Trp/-His/-Ade с добавлением X-α-gal синие колонии дрожжей присутствовали при трансформации в pGBKT7-MdPIF или pGBKT7-MdMYB23, тогда как отрицательный контроль ничего не показывал. Эти результаты показали, что белки MdPIF характеризуются транскрипционной активностью в дрожжевых клетках.
2.6. Анализ экспрессии MdPIF в ответ на абиотические стрессы и гормоны
На основании анализа цис-действующих элементов в промоторах генов MdPIF (и таблицы S2) было разумно предположить, что MdPIF участвуют в ответе на Абиотические факторы и растительные гормоны. Чтобы проверить нашу гипотезу, мы затем исследовали паттерны экспрессии четырех репрезентативных генов MdPIF ( MdPIF1 , MdPIF3 , MdPIF4 и MdPIF8 ) после обработки 100 мМ солевым стрессом (который имитировал солевой стресс) , 8% ПЭГ 6000 (который имитировал стресс от засухи), 100 мкМ индол-3-уксусной кислоты (ИУК), 100 мкМ АБК и 100 мкМ GA соответственно.
Как показано на рисунке , уровень экспрессии генов MdPIF в экспериментальных группах указывает на чувствительность MdPIF к нескольким абиотическим стрессам и гормонам, несмотря на колебания экспрессии MdPIF в контрольной группе. В частности, при обработке NaCl уровни экспрессии всех генов MdPIF были значительно снижены через 6 часов по сравнению с контролем, что свидетельствует о том, что солевой стресс подавлял транскрипцию генов MdPIF и что подавленная экспрессия генов MdPIFs может быть вовлечен в устойчивость к солевому стрессу (а).Кроме того, при обработке ПЭГ 6000 уровни экспрессии MdPIF1 и MdPIF3 значительно увеличивались через 12 часов, тогда как уровни экспрессии MdPIF4 и MdPIF8 резко повышались на ранней стадии (1-часовой и 2-часовой баллы соответственно). Однако, когда уровни экспрессии MdPIF8 снижались после пика (2 часа), уровни экспрессии MdPIF4 оставались на высоком уровне и достигли пика через 6 часов (b).
Анализ экспрессии генов MdPIF в ответ на абиотические стрессоры и гормоны.Уровни экспрессии MdPIF в ответ на NaCl ( a ), PEG 6000 ( b ), индол-3-уксусную кислоту (IAA) ( c ), ABA ( d ) , , 2 Г.А. ( и ). Данные представляют собой среднее значение ± стандартное отклонение для трех независимых повторов. Звездочками отмечены значимые отличия от контроля (*, p < 0,05; **, p < 0,01).
Помимо реакции на абиотические стрессы, уровни экспрессии генов MdPIF также в разной степени подвергались влиянию экзогенных гормонов.Например, при обработке ИУК уровни экспрессии всех генов MdPIF снижались через 3 часа, а затем постепенно увеличивались с 6 до 12 часов (с). При обработке ABA уровни экспрессии MdPIF3 и MdPIF4 были высоко экспрессированы через 12 часов по сравнению с контролем, тогда как уровни экспрессии MdPIF1 и MdPIF8 повышались как через 1 час, так и через 12 часов (d).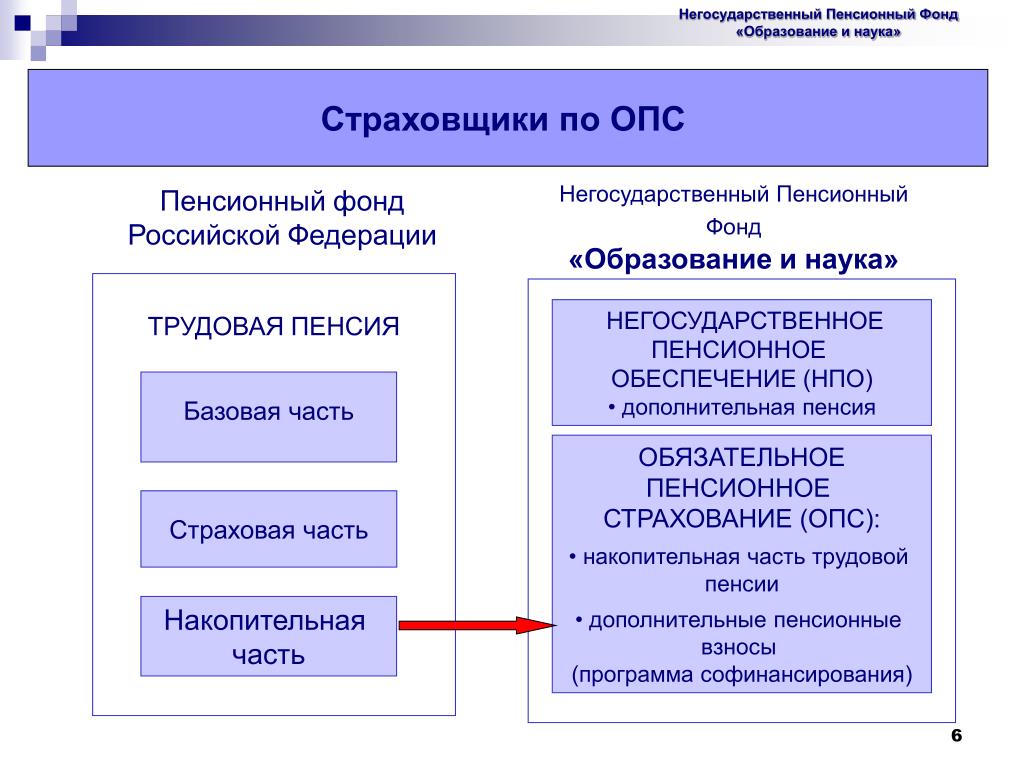 Кроме того, MdPIF1, MdPIF3, и MdPIF4 показали значительное повышение уровня транскриптов через 3 часа после обработки ГА, тогда как MdPIF8 был сильно экспрессирован через 12 часов (e).В совокупности эти данные свидетельствуют о том, что гены яблока MdPIF широко вовлечены во множественные реакции на абиотический стресс и гормоны.
Кроме того, MdPIF1, MdPIF3, и MdPIF4 показали значительное повышение уровня транскриптов через 3 часа после обработки ГА, тогда как MdPIF8 был сильно экспрессирован через 12 часов (e).В совокупности эти данные свидетельствуют о том, что гены яблока MdPIF широко вовлечены во множественные реакции на абиотический стресс и гормоны.
2.7. Сверхэкспрессия MdPIF4 снижает чувствительность к ABA в мозолях яблони
Среди семи промоторов гена MdPIF было обнаружено большое количество ABA-чувствительных элементов, что указывает на то, что эти гены Apple PIF могут быть вовлечены в реакцию ABA. Промотор MdPIF4 содержал наибольшее количество АБК-чувствительных элементов (), а на уровень экспрессии MdPIF4 значительно влияла обработка АБК (d).Поэтому мы предположили, что MdPIF4 с высокой вероятностью будет участвовать в сигнальных путях ABA. Сначала мы получили две трансгенные линии каллусов яблони со сверхэкспрессией MdPIF4 .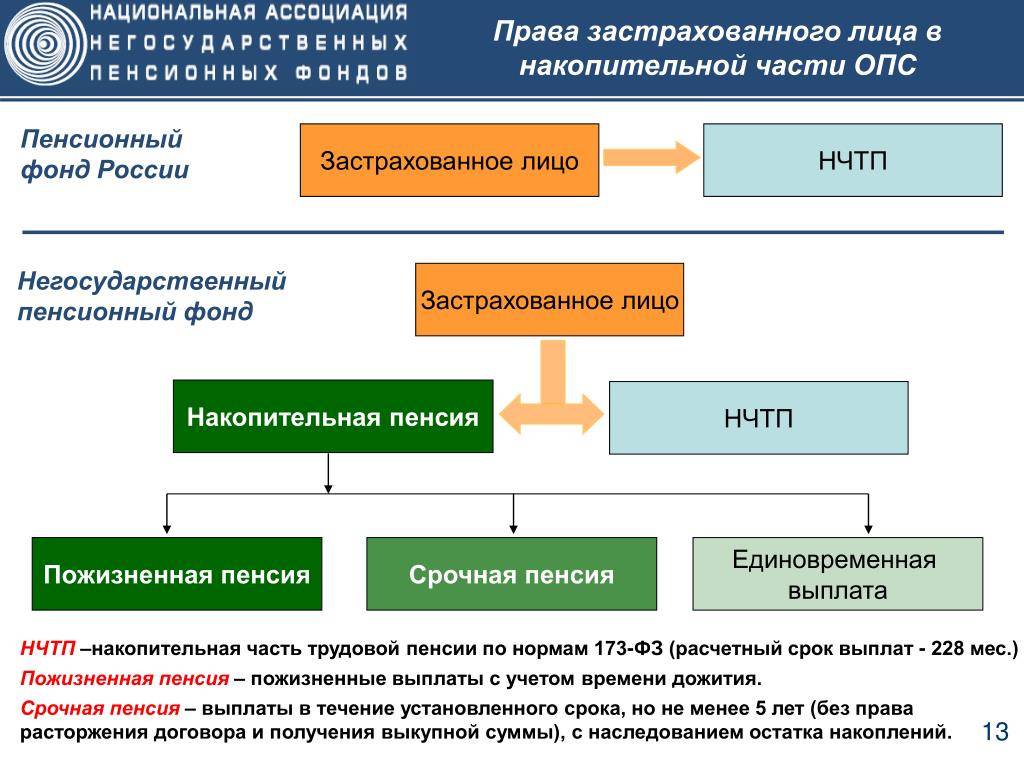 Трансгенные каллюсы генерировали гораздо более высокие уровни транскрипции MdPIF4 по сравнению с контролем дикого типа, что свидетельствует о том, что MdPIF4 успешно трансформировался в каллюсе (а). При имитационной обработке рост WT и каллюса MdPIF4-OE был согласованным, и они имели сходную массу в свежем виде, а также содержание MDA, который является наиболее мутагенным продуктом перекисного окисления липидов, и его содержание было разработано для важный показатель для измерения степени повреждения растений, вызванного стрессами [39,40,41] (b,d).Экзогенная обработка ABA ограничивает удлинение клеток и, таким образом, ингибирует рост каллуса яблони, что было продемонстрировано в предыдущих отчетах [42]. При добавлении АБК в среду рост каллусов WT и MdPIF4 -OE подавлялся по сравнению с таковым в имитационных условиях, а более высокие концентрации АБК ухудшали его (б). Тем не менее, каллус MdPIF4 -OE рос намного лучше, чем WT, что свидетельствует о том, что сверхэкспрессия MdPIF4 в каллюсе яблони снижает их чувствительность к АБК (b).
Трансгенные каллюсы генерировали гораздо более высокие уровни транскрипции MdPIF4 по сравнению с контролем дикого типа, что свидетельствует о том, что MdPIF4 успешно трансформировался в каллюсе (а). При имитационной обработке рост WT и каллюса MdPIF4-OE был согласованным, и они имели сходную массу в свежем виде, а также содержание MDA, который является наиболее мутагенным продуктом перекисного окисления липидов, и его содержание было разработано для важный показатель для измерения степени повреждения растений, вызванного стрессами [39,40,41] (b,d).Экзогенная обработка ABA ограничивает удлинение клеток и, таким образом, ингибирует рост каллуса яблони, что было продемонстрировано в предыдущих отчетах [42]. При добавлении АБК в среду рост каллусов WT и MdPIF4 -OE подавлялся по сравнению с таковым в имитационных условиях, а более высокие концентрации АБК ухудшали его (б). Тем не менее, каллус MdPIF4 -OE рос намного лучше, чем WT, что свидетельствует о том, что сверхэкспрессия MdPIF4 в каллюсе яблони снижает их чувствительность к АБК (b). Свежий вес каллусов яблони со сверхэкспрессией MdPIF4 был значительно выше, чем у WT при обработке АБК (с), что согласуется с фенотипическими результатами, показанными на b. Более того, каллюс MdPIF4-OE имел более низкое содержание MDA по сравнению с таковым у WT, что позволяет предположить, что сверхэкспрессия MdPIF4 облегчала повреждение, вызванное ABA, и, таким образом, более эффективно способствовала росту каллюса при обработке ABA.
Свежий вес каллусов яблони со сверхэкспрессией MdPIF4 был значительно выше, чем у WT при обработке АБК (с), что согласуется с фенотипическими результатами, показанными на b. Более того, каллюс MdPIF4-OE имел более низкое содержание MDA по сравнению с таковым у WT, что позволяет предположить, что сверхэкспрессия MdPIF4 облегчала повреждение, вызванное ABA, и, таким образом, более эффективно способствовала росту каллюса при обработке ABA.
Пониженная чувствительность АБК каллуса трансгенного яблока MdPIF4 .( a ) Анализ экспрессии MdPIF4 в WT и трансгенных каллюсах. В качестве внутреннего контроля использовали 18S рибосомную РНК. ( b ) Фенотипы 14-дневных каллусов трансгенных яблок и дикого типа, культивируемых на среде Мурасиге и Скуга (МС), содержащей различные концентрации АБК, в течение 20 дней. ( c ) Статистический анализ свежего веса каллюсов трансгенных яблок и яблок дикого типа после обработки.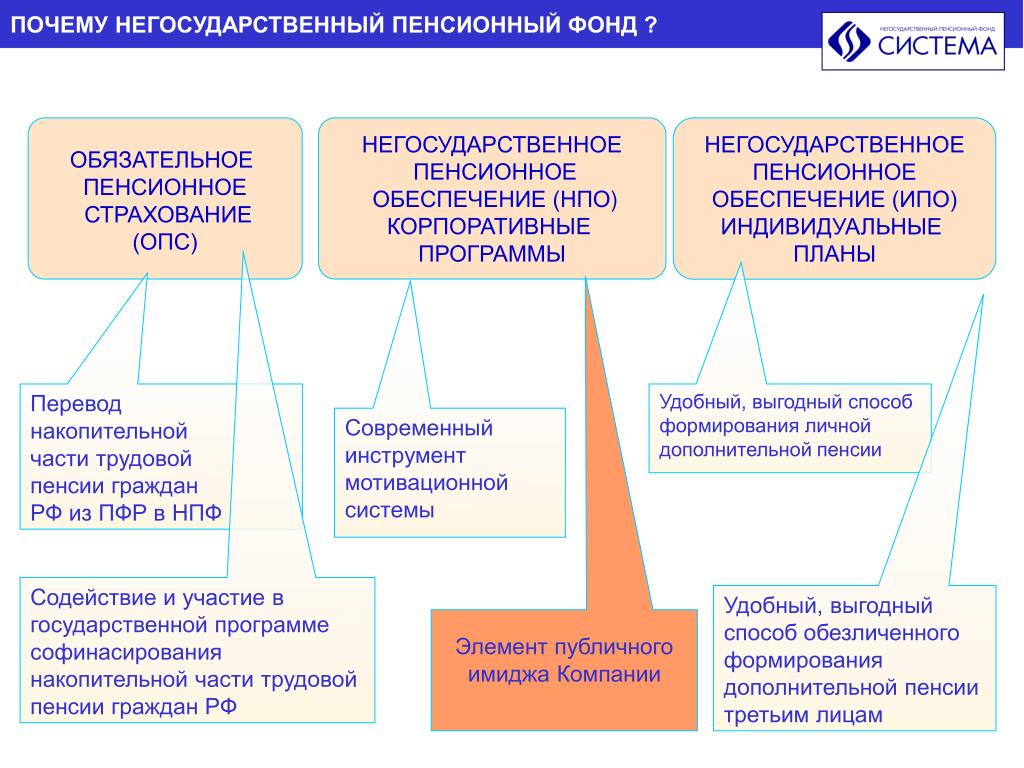 ( d ) Статистический анализ концентраций MDA в каллюсах трансгенных и WT яблок после обработки.( e ) Экспрессия генов генов ответа ABA в WT и трансгенных каллюсах после обработки ABA. Данные представляют собой среднее значение ± стандартное отклонение для трех независимых повторов. Звездочками отмечены значимые отличия от контроля (*, p < 0,05; **, p < 0,01).
( d ) Статистический анализ концентраций MDA в каллюсах трансгенных и WT яблок после обработки.( e ) Экспрессия генов генов ответа ABA в WT и трансгенных каллюсах после обработки ABA. Данные представляют собой среднее значение ± стандартное отклонение для трех независимых повторов. Звездочками отмечены значимые отличия от контроля (*, p < 0,05; **, p < 0,01).
Для дальнейшего изучения роли MdPIF4 в регуляции ответа ABA мы протестировали характер экспрессии нескольких генов ответа ABA в каллюсах WT и трансгенных яблок. Гены ответа ABA, такие как MdEM1 , MdEM6 , MdRAB18 и MdRD29A , которые, как сообщалось, участвуют в нижестоящих сигнальных путях ABA, а также MdAREB3.1, , который регулирует сигнальные пути АБК [43], были значительно снижены в каллусе MdPIF4 — OE по сравнению с таковым в яблоках WT (e), что позволяет предположить, что MdPIF4 может участвовать в модуляции экспрессии ответа АБК. гены, регулирующие чувствительность к АБК.
гены, регулирующие чувствительность к АБК.
3. Обсуждение
После завершения полногеномного секвенирования яблок [44,45,46] было идентифицировано множество семейств транскрипционных факторов, таких как MYB [47], WRKY [48], BBX [49] и бГЛГ [32].Некоторые из основных генов были охарактеризованы, и их биологические функции широко изучены, например, MYB1 [50], WRKY40 [51] и BBX22 [52]. Хотя PIF играют важную роль в росте и развитии растений, систематическая характеристика и функциональное исследование семейства PIF у яблони не проводились, за исключением MdPIF1 [31]. Здесь был проведен систематический анализ семейства генов MdPIF , который дает направления для дальнейших исследований гена PIF у яблок и отбора генов важных признаков.
3.1. PIF являются транскрипционными факторами, консервативными среди различных видов растений
В этом исследовании семь генов MdPIF были идентифицированы у яблок, что аналогично числам, указанным для других растений: восемь для Arabidopsis [9], шесть для риса [53]. ] и семь для кукурузы [38]. Филогенетический анализ показал, что MdPIF были разделены на четыре клады (а). Результаты отличаются от предыдущих сообщений об эволюционных отношениях белков PIF у кукурузы и риса, в которых белки PIF сгруппированы в три клады [38,53].Анализ последовательностей белка MdPIF показал, что все MdPIF имеют общий консервативный домен bHLH и мотив APB. Пять MdPIF (MdPIF1, MdPIF2, MdPIF3, MdPIF4 и MdPIF5) имеют мотив APA, сходный с AtPIF1 и AtPIF3, что позволяет предположить, что эти MdPIF могут быть близки к AtPIF1 и AtPIF3 в эволюционном масштабе (b). Кроме того, стоит отметить, что MdPIF7 и MdPIF8 обнаруживают аминокислотную замену в домене APB, которая изменяет консервативный остаток Q на X и E соответственно (рис. S3a) [14].Между тем, добавление аминокислотного остатка в домен bHLH было специфичным в MdPIF1 и MdPIF2 по сравнению с другими MdPIF, что позволяет предположить, что они могут иметь потенциально уникальную функцию (рис. S3a). Анализ паттерна экспрессии в различных тканях яблони показал, что все генов MdPIF были конститутивно экспрессированы в исследованных тканях с относительно высокими уровнями в листьях (a-g), что сходно с паттерном экспрессии AtPIF и ZmPIF [4].
] и семь для кукурузы [38]. Филогенетический анализ показал, что MdPIF были разделены на четыре клады (а). Результаты отличаются от предыдущих сообщений об эволюционных отношениях белков PIF у кукурузы и риса, в которых белки PIF сгруппированы в три клады [38,53].Анализ последовательностей белка MdPIF показал, что все MdPIF имеют общий консервативный домен bHLH и мотив APB. Пять MdPIF (MdPIF1, MdPIF2, MdPIF3, MdPIF4 и MdPIF5) имеют мотив APA, сходный с AtPIF1 и AtPIF3, что позволяет предположить, что эти MdPIF могут быть близки к AtPIF1 и AtPIF3 в эволюционном масштабе (b). Кроме того, стоит отметить, что MdPIF7 и MdPIF8 обнаруживают аминокислотную замену в домене APB, которая изменяет консервативный остаток Q на X и E соответственно (рис. S3a) [14].Между тем, добавление аминокислотного остатка в домен bHLH было специфичным в MdPIF1 и MdPIF2 по сравнению с другими MdPIF, что позволяет предположить, что они могут иметь потенциально уникальную функцию (рис. S3a). Анализ паттерна экспрессии в различных тканях яблони показал, что все генов MdPIF были конститутивно экспрессированы в исследованных тканях с относительно высокими уровнями в листьях (a-g), что сходно с паттерном экспрессии AtPIF и ZmPIF [4]. 38,54]. Ядерная локализация (a) и активность активации транскрипции (c) MdPIFs указывают на то, что они являются типичными факторами транскрипции и что они могут функционировать как позитивные регуляторы нижестоящей экспрессии генов.
38,54]. Ядерная локализация (a) и активность активации транскрипции (c) MdPIFs указывают на то, что они являются типичными факторами транскрипции и что они могут функционировать как позитивные регуляторы нижестоящей экспрессии генов.
3.2. PIF действуют как молекулярный концентратор в интеграции экологических и гормональных сигнальных путей
Помимо того, что они являются негативными регуляторами фотоморфогенеза, растущее количество данных указывает на то, что PIF действуют как сигнальный концентратор и играют ключевые роли во многих процессах, включая синтез антоцианов [18]. ]; устойчивость к засухе [22,23], соли [23] и холоду [55]; сигнальные пути растительных гормонов (ГК, БР, ауксин) [56]; и даже в регуляции иммунитета растений [57].
Цис-действующие элементы в области промотора гена могут распознаваться несколькими клеточными белками, такими как факторы транскрипции, для регуляции транскрипции и экспрессии генов в ответ на сигналы окружающей среды и гормоны. Например, экспрессия мРНК AtPIF4 активируется высокой температурой. Далее было обнаружено, что высокая температура инактивирует AtELF3 (раннее цветение 3), который подавляет экспрессию AtPIF4 путем прямого связывания со своим промотором [56]. Кроме того, аффинность связывания AtELF3 с промотором AtPIF4 снижается при высокой температуре (27 °C) по сравнению с таковой при нормальной температуре (22 °C) [58]. При анализе промотора генов MdPIF оказалось, что светочувствительные родственные элементы значительно преобладают (и таблица S2).Объединив изменения уровня транскрипции генов MdPIF днем и ночью в ч, мы обнаружили, что генов MdPIF регулируются светоопосредованным суточным ритмом. В дополнение к фотопериоду было предсказано, что множественные цис-действующие элементы реагируют на растительные гормоны (АБК, ГА и ЖАК) и абиотические стрессы (засуха, низкая температура и раны) (и Таблица S2). Предыдущие исследования показали, что некоторые гормоны, такие как АБК, ГК и ЖАК, играют ключевую роль в регуляции реакции растений на стрессовые условия [59].
Например, экспрессия мРНК AtPIF4 активируется высокой температурой. Далее было обнаружено, что высокая температура инактивирует AtELF3 (раннее цветение 3), который подавляет экспрессию AtPIF4 путем прямого связывания со своим промотором [56]. Кроме того, аффинность связывания AtELF3 с промотором AtPIF4 снижается при высокой температуре (27 °C) по сравнению с таковой при нормальной температуре (22 °C) [58]. При анализе промотора генов MdPIF оказалось, что светочувствительные родственные элементы значительно преобладают (и таблица S2).Объединив изменения уровня транскрипции генов MdPIF днем и ночью в ч, мы обнаружили, что генов MdPIF регулируются светоопосредованным суточным ритмом. В дополнение к фотопериоду было предсказано, что множественные цис-действующие элементы реагируют на растительные гормоны (АБК, ГА и ЖАК) и абиотические стрессы (засуха, низкая температура и раны) (и Таблица S2). Предыдущие исследования показали, что некоторые гормоны, такие как АБК, ГК и ЖАК, играют ключевую роль в регуляции реакции растений на стрессовые условия [59].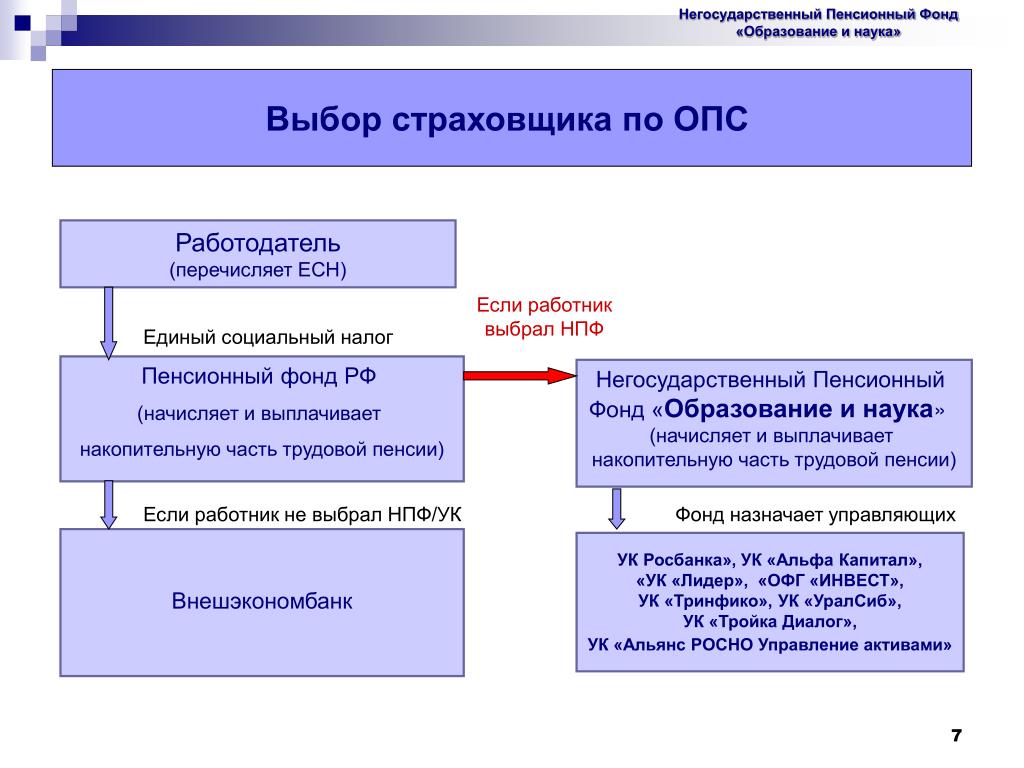 Наши данные также подтвердили, что экспрессия MdPIF может индуцироваться несколькими абиотическими стрессами и гормонами (4). Например, MdPIF1 и MdPIF4 значительно усиливались засухой и обработкой GA (b, e). Было предсказано, что мотивы, участвующие в реакции на засуху (MBS: CAACTG) и ABA (ABRE: ACGTG), присутствуют в промоторе MdPIF3 (и таблица S2), а уровень экспрессии MdPIF3 значительно индуцируется. после засухи и обработки АБК в течение 12 ч (б, г).Кроме того, цис-элементы, которые функционируют в ответ на ГА, присутствовали в промоторах MdPIF3 и MdPIF4 , и оба они были индуцированы для экспрессии в различной степени обработкой ГА (e). Кроме того, в промоторах большинства генов MdPIF были обнаружены мотивы, чувствительные к АБК, и эти гены в разной степени индуцировались при обработке АБК (d). Эти данные свидетельствуют о том, что цис-элементы в промоторной области MdPIF могут быть функциональными в их ответе на абиотические стрессы и гормоны.
Наши данные также подтвердили, что экспрессия MdPIF может индуцироваться несколькими абиотическими стрессами и гормонами (4). Например, MdPIF1 и MdPIF4 значительно усиливались засухой и обработкой GA (b, e). Было предсказано, что мотивы, участвующие в реакции на засуху (MBS: CAACTG) и ABA (ABRE: ACGTG), присутствуют в промоторе MdPIF3 (и таблица S2), а уровень экспрессии MdPIF3 значительно индуцируется. после засухи и обработки АБК в течение 12 ч (б, г).Кроме того, цис-элементы, которые функционируют в ответ на ГА, присутствовали в промоторах MdPIF3 и MdPIF4 , и оба они были индуцированы для экспрессии в различной степени обработкой ГА (e). Кроме того, в промоторах большинства генов MdPIF были обнаружены мотивы, чувствительные к АБК, и эти гены в разной степени индуцировались при обработке АБК (d). Эти данные свидетельствуют о том, что цис-элементы в промоторной области MdPIF могут быть функциональными в их ответе на абиотические стрессы и гормоны.
Растущий объем данных указывает на то, что PIF4 действует как ключевой интегратор множественных сигнальных путей [56], включая световые (скомоторфогенез) и температурные сигнальные пути [21], гормональные сигнальные пути (ауксин, GA и BR), и выходные пути циркадных часов. Здесь мы обнаружили, что обработка АБК влияла на экспрессию MdPIF4 (d), а сверхэкспрессия MdPIF4 снижала чувствительность каллусов яблони к АБК за счет подавления экспрессии генов, чувствительных к АБК (), что указывает на то, что MdPIF4 может играть роль отрицательную регуляторную роль в сигнальных путях АБК, о которой еще не сообщалось, и механизм, лежащий в основе этого, все еще нуждается в дальнейшем изучении.Кроме того, у яблок идентифицировано несколько генов, участвующих в реакции АБК. Экспрессия семи генов MdAGO из семейства MdAGO была значительно усилена обработкой АБК [60]. Сверхэкспрессия MdSINA2 повышала чувствительность к АБК в каллусе яблони и Arabidopsis [61].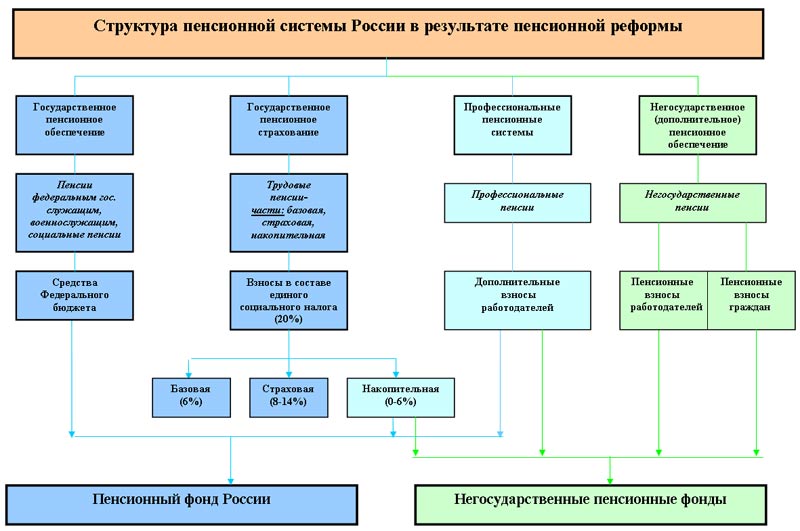 Хотя регуляторный механизм еще предстоит решить, идентификация этих ABA-чувствительных генов обогатила наши знания о сети сигнальных путей ABA у яблок и послужит основой для будущих исследований их функций.
Хотя регуляторный механизм еще предстоит решить, идентификация этих ABA-чувствительных генов обогатила наши знания о сети сигнальных путей ABA у яблок и послужит основой для будущих исследований их функций.
4. Материалы и методы
4.1. Растительные материалы и условия роста
Образцы ткани яблони ( Malus domestica ‘Royal Gala’) (стебель, молодой корень, зрелый лист, цветок и созревающий плод) были собраны с трех 10-летних яблонь в искусственно управляемом orchard для исследования паттерна экспрессии MdPIF s [62,63]. Следует отметить, что созревающие плоды были собраны через 150 дней после цветения, а корни были взяты из молодых корней, которые были недавно выращены в этом году.Проростки тканевой культуры яблони ( Malus domestica ‘Royal Gala’) поддерживали на среде Мурасиге и Скуга (MS) с 0,5 мМ 6-бензиламинопурином (6-BA) и 0,1 мМ индол-3-уксусной кислотой (IAA) в течение 16 ч/8 ч светового/темнового фотопериода (интенсивность света 70 мкмоль м -2 с -1 ) в культуральной комнате при 25 °C [32]. Через 21 день роста проростки переносили в среду, индуцирующую рост корней (среда MS в пределах 0,1 мМ ИУК), для индукции роста корней [64]. Эти саженцы яблони, выращенные в тканях, использовали для измерения дневной экспрессии в 8:00, 10:00, 12:00, 14:00, 16:00, 18:00, 20:00, 22:00, 24:00, 2 :00 2д (второй день), 4:00 2д , 6:00 2д и 8:00 2д .Свет начинался с 8 часов и заканчивался в 24 часа, после чего наступали 8 часов темноты. Для обработки от абиотических стрессов необработанные саженцы яблони переводили в водные условия культивирования. В условиях нормального фотопериода проростки с одинаковым состоянием роста обрабатывали NaCl (100 мМ), полиэтиленгликолем 6000 (ПЭГ 6000, 8%), ИУК (100 мкМ), АБК (100 мкМ) и гиббереллином (ГА) ( 100 мкМ) соответственно [65,66]. Эти проростки собирали через 0 ч, 1 ч, 2 ч, 3 ч, 6 ч и 12 ч после обработки, немедленно замораживали жидким азотом и хранили при -80 °C для дальнейшего анализа [67].
Через 21 день роста проростки переносили в среду, индуцирующую рост корней (среда MS в пределах 0,1 мМ ИУК), для индукции роста корней [64]. Эти саженцы яблони, выращенные в тканях, использовали для измерения дневной экспрессии в 8:00, 10:00, 12:00, 14:00, 16:00, 18:00, 20:00, 22:00, 24:00, 2 :00 2д (второй день), 4:00 2д , 6:00 2д и 8:00 2д .Свет начинался с 8 часов и заканчивался в 24 часа, после чего наступали 8 часов темноты. Для обработки от абиотических стрессов необработанные саженцы яблони переводили в водные условия культивирования. В условиях нормального фотопериода проростки с одинаковым состоянием роста обрабатывали NaCl (100 мМ), полиэтиленгликолем 6000 (ПЭГ 6000, 8%), ИУК (100 мкМ), АБК (100 мкМ) и гиббереллином (ГА) ( 100 мкМ) соответственно [65,66]. Эти проростки собирали через 0 ч, 1 ч, 2 ч, 3 ч, 6 ч и 12 ч после обработки, немедленно замораживали жидким азотом и хранили при -80 °C для дальнейшего анализа [67].
Каллусы яблока сорта «Орин» ( Golden Delicious × Indo ) культивировали на среде MS, содержащей 1,5 мг/л 2,4-дихлорфеноксиуксусной кислоты (2, 4-D) и 0,4 мг/л 6-BA при 24 °С в течение 20 дней в темноте [52].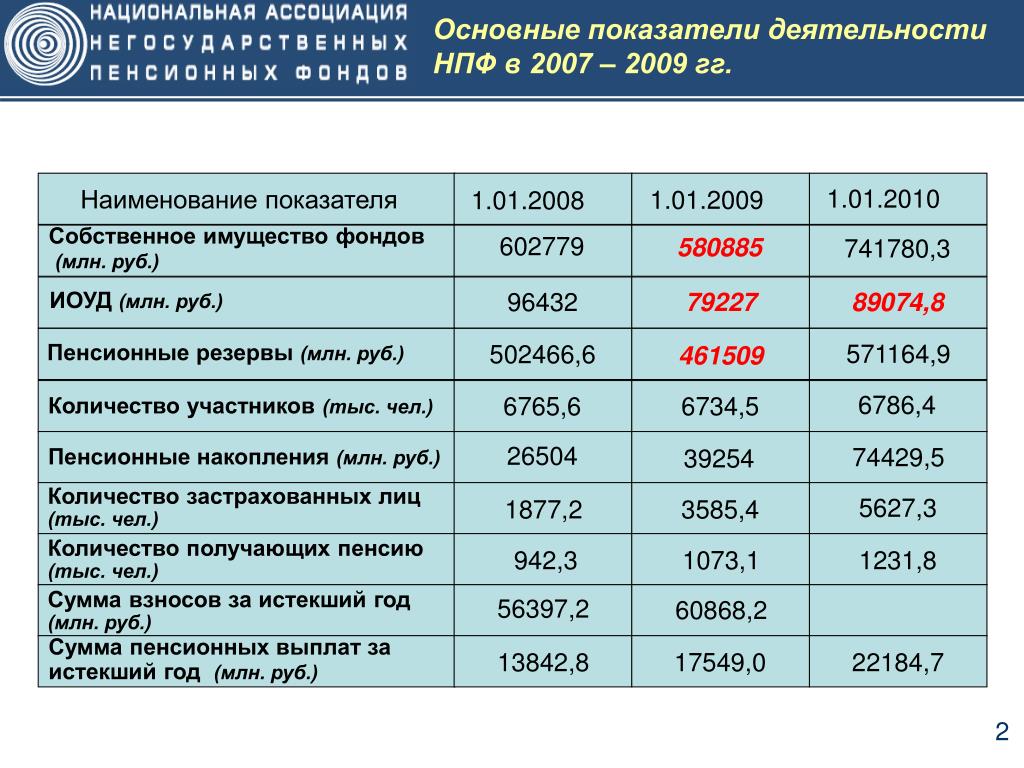
4.2. Конструирование плазмиды и получение трансгенного растительного материала
Полноразмерную кДНК MdPIF4 клонировали в вектор pRI 101-AN (Takara, Далянь, Китай) для получения конструкции сверхэкспрессии MdPIF4 .Затем пустой вектор pRI 101-AN и рекомбинантную конструкцию трансформировали в штамм Agrobacterium tumefaciens LBA4404. Эти агробактерии выращивали в среде лизогенного бульона (LB) с добавлением 50 мг/мл канамицина и 50 мг/мл рифампицина [68].
Трансгенные каллусы яблони были получены путем трансформации, опосредованной агробактериями [69]. Вкратце, 14-дневные каллусы яблони совместно культивировали со штаммами агробактерий, несущими пустой вектор или рекомбинантные конструкции, в темноте в течение 20 мин.Затем трансформированные каллусы яблони высевали на селективную среду с добавлением 250 мг/л карбенициллина и 30 мг/л канамицина [70]. Успешно трансгенные яблочные мозоли выросли примерно за четыре недели.
4.
 3. Идентификация, хромосомное расположение и функциональная аннотация генов MdPIF
3. Идентификация, хромосомное расположение и функциональная аннотация генов MdPIFДля идентификации генов MdPIF из генома Malus domestica мы загрузили восемь пептидных последовательностей PIF из базы данных Arabidopsis [71].Затем был проведен поиск MdPIF в базе данных генома яблони ( Malus domestica v1.0) с использованием белковых последовательностей Arabidopsis [72]. Были отобраны восемь белковых последовательностей-кандидатов, и затем мы использовали базу данных SMART [73], чтобы проверить, содержат ли эти кандидаты консервативные домены bHLH и APB.
Хромосомное расположение этих генов было найдено в файле аннотации генома яблони ( Malus domestica .v1.0.consensus.gff) из Базы данных геномов розоцветных (GDR) [74]. генов MdPIF были отправлены в онлайн-программу MG2C [75] для картирования хромосом генов. Молекулярная масса, теоретическая изоэлектрическая точка (pI) и гидрофобность белка этих MdPIF были проанализированы с использованием онлайн-инструмента ProtParam [76].
4.4. Филогенетический анализ белков PIF
Последовательности белков AtPIF были получены из базы данных Arabidopsis [71,77]. Белковые последовательности PIF риса были получены из проекта аннотации риса [78].На основе всех последовательностей белков PIF из Arabidopsis , риса и яблока было построено филогенетическое дерево с использованием программы MEGA5.1 методом соседнего соединения со следующими настройками: метод Bootstrap, 1000 повторов и модель Пуассона [79, 80].
4.5. Структура гена (интрон/экзон), множественное выравнивание последовательностей и анализ консервативных мотивов
Файл базы данных генома яблони ( Malus domestica .v1.0.consensus.gff) был загружен из базы данных GDR.Информация о структуре генов MdPIF была получена с использованием сервера визуализации признаков генов GSDS 2.0 [81,82,83]. Выравнивание последовательностей выполняли с использованием программного обеспечения DANMAN с параметрами по умолчанию [42,84]. Анализ консервативных мотивов проводили с помощью программы MEME [85], а для отображения структуры мотива использовали TBtools [86].
Анализ консервативных мотивов проводили с помощью программы MEME [85], а для отображения структуры мотива использовали TBtools [86].
4.6. Анализ цис-действующих элементов в промоутерах MdPIF
Фрагмент длиной 1500 п.н. перед стартовым кодоном генов MdPIF считался промоторной областью, и последовательности промоторов были отправлены в PlantCARE [87] для анализа цис-действующие элементы [88,89].
4.7. Экстракция РНК и количественный анализ ПЦР в реальном времени (qRT-PCR)
Всего РНК экстрагировали с помощью набора реагентов RNAplant Plus (Tiangen, Пекин, Китай). Обратную транскрипцию выполняли с использованием набора реагентов PrimeScript™ RT (Takara, Shiga, Japan). Реакционная система для qRT-PCR содержала 10 мкл премикса SYBR Ex Taq™ (Takara), 0,8 мкл (10 мМ) каждого праймера и 50 нг кДНК в общем объеме 20 мкл. Процедуру реакции qRT-PCR проводили в соответствии с методами, о которых сообщалось ранее [90].Экспериментальный образец ПЦР имел три повторения, а 18S рибосомная РНК служила контролем [91].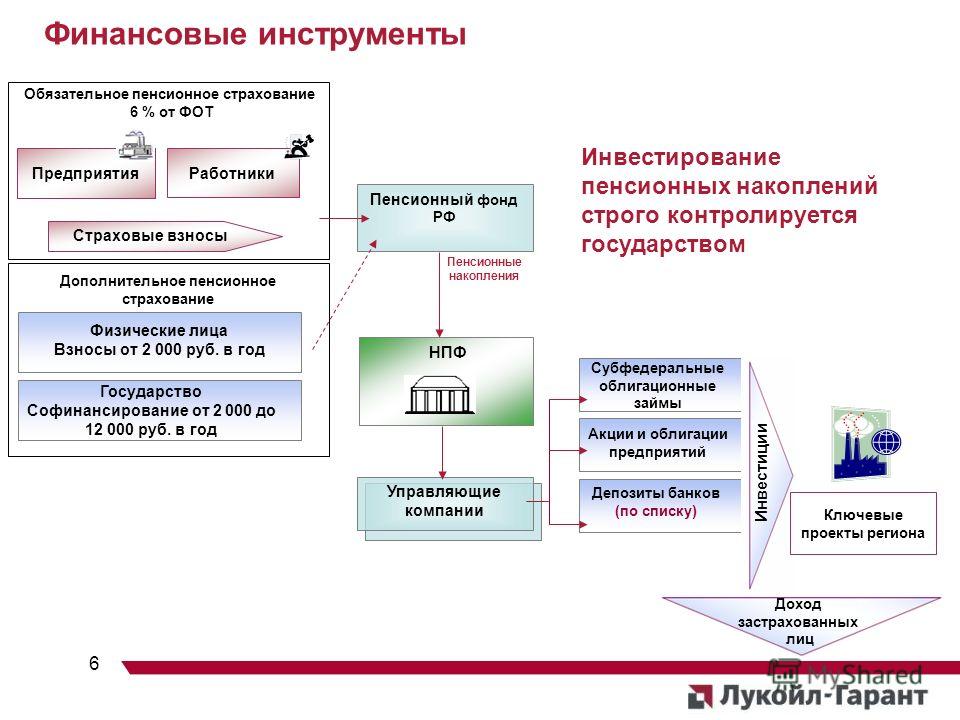 Для анализа данных использовали метод 2 −ΔΔCT 2 −ΔΔCT . Все праймеры, использованные в этом исследовании, показаны в таблице S1.
Для анализа данных использовали метод 2 −ΔΔCT 2 −ΔΔCT . Все праймеры, использованные в этом исследовании, показаны в таблице S1.
4.8. Анализ субклеточной локализации
Кодирующие последовательности MdPIF1 , MdPIF3 , MdPIF4, и MdPIF8 были клонированы в вектор 35S::GFP для трансляцииРекомбинантные векторы экспрессии 35S::MdPIF-GFP трансформировали в штамм Agrobacterium LBA4400 соответственно. Затем конструкции 35S::MdPIF-GFP , несущие агробактерии, инъецировали в листья Nicotiana benthamiana [92] и культивировали в темноте в течение 2 сут. Сигнал GFP в клетках наблюдали с помощью конфокального микроскопа высокого разрешения (LSCM Meta; Carl Zeiss, Zena, Германия) [93].
4.9. Анализ транскрипционной активности
Кодирующие последовательности MdPIF1 , MdPIF3 , MdPIF4, и MdPIF8 были клонированы в вектор pGBKT7 с использованием метода гомологичной рекомбинации. Сайтами рестрикционных эндонуклеаз были EcoRI и SalI (таблица S1). Поскольку в нашей лаборатории было доказано, что MdMYB23 (номер доступа в GenBank: MDP0000230141) обладает транскрипционной активностью [94], его можно было использовать в качестве положительного контроля. Слитые векторы pGBKT7-MdPIF с использованием вектора pGBKT7 (отрицательный контроль) и pGBKT7-MdMYB23 (положительный контроль) переносили в штамм дрожжей Ah209. Затем их культивировали на среде SD (SD/-Trp, SD/-Trp/-His/-Ade/X-α-gal) при 28 °С в течение 3–5 сут.
Сайтами рестрикционных эндонуклеаз были EcoRI и SalI (таблица S1). Поскольку в нашей лаборатории было доказано, что MdMYB23 (номер доступа в GenBank: MDP0000230141) обладает транскрипционной активностью [94], его можно было использовать в качестве положительного контроля. Слитые векторы pGBKT7-MdPIF с использованием вектора pGBKT7 (отрицательный контроль) и pGBKT7-MdMYB23 (положительный контроль) переносили в штамм дрожжей Ah209. Затем их культивировали на среде SD (SD/-Trp, SD/-Trp/-His/-Ade/X-α-gal) при 28 °С в течение 3–5 сут.
4.10. Обработка каллюсов яблони и физиологические измерения
14-дневные каллюсы дикого типа и трансгенные яблочные каллюсы культивировали на среде MS, содержащей 0 мкМ, 50 мкМ и 100 мкМ АБК. Электронные весы (одна тысячная) использовали для измерения веса яблочных каллюсов для определения роста каллюсов. Содержание малонового диальдегида (МДА) измеряли с использованием метода тиобарбитуровой кислоты, о котором сообщалось ранее [43].
4.
 11. Статистический анализ
11. Статистический анализВсе данные были получены из трех независимых экспериментов с тремя биологическими дубликатами в каждом эксперименте.Программное обеспечение DPS использовалось для анализа значимой разницы, и значение p <0,05 считалось значимой разницей [95]. Столбики погрешностей представляют собой стандартное отклонение.
(PDF) Транскрипционная активность Pygopus усиливается за счет его взаимодействия с белком, связывающим цАМФ-ответный элемент (CREB)-связывающим белком
Pygopus-зависимая передача сигналов Wnt 501
16 Shahbazian, MD and Grunstein, M. ( 2007) Функции сайт-специфического ацетилирования гистонов
и деацетилирования.Анну. Преподобный Биохим. 76, 75–100
17 Фидлер М., Санчес-Баррена М.Дж., Некрасов М., Мещанек Дж., Рыбин В., Мюллер Дж.,
Эванс П. и Бинц М. (2008)Расшифровка хвоста метилированного гистона h4 сигнальным комплексом Pygo-BCL9
Wnt. Мол. Cell 30, 507–518
18 Feng, Y., Lee, N. and Fearon, ER (2003) TIP49 регулирует β-катенин-опосредованную неопластическую трансформацию
and Fearon, ER (2003) TIP49 регулирует β-катенин-опосредованную неопластическую трансформацию
и индукцию гена-мишени Т-клеточного фактора посредством воздействия на хроматин
реконструкция.Рак рез. 63, 8726–8734
19 Parker, D.S., Ni, Y.Y., Chang, J.L., Li, J. and Cadigan, K.M. (2008) Передача сигналов Wingless
индуцирует широко распространенное ремоделирование хроматина локусов-мишеней. Мол. Клеточная биол. 28,
1815–1828
20 Sustmann, C., Flach, H., Ebert, H., Eastman, Q. and Grosschedl, R. (2008)
Специфическая для типа клеток функция BCL9 включает транскрипционную домен активации, который
взаимодействует с β-катенином. Мол. Клеточная биол. 28, 3526–3537
21 Баркер, Н., Hurlstone, A., Musisi, H., Miles, A., Bienz, M. and Clevers, H. (2001) Фактор ремоделирования хроматина
Brg-1 взаимодействует с β-катенином, способствуя активации целевого гена
. EMBO J. 20, 4935–4943
22 Takemaru, K.I. and Moon, R.T. (2000) Коактиватор транскрипции CBP взаимодействует с
and Moon, R.T. (2000) Коактиватор транскрипции CBP взаимодействует с
β-катенином для активации экспрессии генов. Дж. Клеточная биология. 149, 249–254
23 Hecht, A., Vleminckx, K., Stemmler, MP, van Roy, F. and Kemler, R. (2000) Ацетилтрансферазы
p300/CBP функционируют как коактиваторы транскрипции β-катенина. в
позвоночных.EMBO J. 19, 1839–1850
24 Наир, М., Нагамори, И., Сун, П., Мишра, Д.П., Реом, К., Ли, Б., Сассоне-Корси, П. и
Дай , X. (2008) Ядерный регулятор Pygo2 контролирует спермиогенез и ацетилирование гистонов h4
. Дев. биол. 15, 446–455
25 Эндрюс, П. Г., Кеннеди, М. В., Попадюк, С. М. и Као, К. Р. (2008) Онкогенная
активация промотора Pygopus2 человека с помощью E74-подобного фактора-1. Мол. Рак рез. 6,
259–266
26 Попадюк К.M., Xiong, J., Wells, MG, Andrews, PG, Dankwa, K., Hirasawa, K., Lake,
BB and Kao, KR (2006) Антисмысловое подавление pygopus2 приводит к остановке роста
эпителиальных рак яичников. клин. Рак рез. 12, 2216-2223
клин. Рак рез. 12, 2216-2223
27 Chrivia, J.C., Kwok, R.P., Lamb, N., Hagiwara, M., Montminy, M.R. and Goodman, R.H.
(1993). Фосфорилированный CREB специфически связывается с ядерным белком CBP. Nature 365,
855–859
28 Блэкмор, Т.M., Mercer, C.F., Paterno, G.D. и Gillespie, L.L. (2008) Транскрипционный кофактор
MIER1-β отрицательно регулирует активность гистон-ацетилтрансферазы
CREB-связывающего белка. БМК. Рез. Примечания 1,68
29 Korzus, E., Torchia, J., Rose, DW, Xu, L., Kurokawa, R., McInerney, EM, Mullen, TM,
Glass, CK and Rosenfeld, MG (1998 ) Специфические требования к факторам транскрипции для
коактиваторов и их ацетилтрансферазных функций.Science 279, 703–707
30 Эндрюс, П. Г., Лейк, Б. Б., Попадюк, К. и Као, К. Р. (2007) Требование Pygopus 2
при раке молочной железы. Междунар. Дж. Онкол. 30, 357–363
31 Дин, З., Гиллеспи, Л.Л., Мерсер, Ф.К. и Патерно, Г.Д. (2004) Домен SANT
MI-ER1 человека взаимодействует с Sp1, препятствуя распознаванию GC-бокса и репрессии
транскрипция от собственного промотора. Дж. Биол. хим. 279, 28009–28016
Дж. Биол. хим. 279, 28009–28016
32 Айт-Си-Али, С., Рамирес, С., Barre, F.X., Dkhissi, F., Magnaghi-Jaulin, L., Girault, J.A.,
Robin, P., Knibiehler, M., Pritchard, L.L., Ducommun, B. et al. (1998) Гистон
Ацетилтрансферазная активность СВР контролируется циклозависимыми киназами и
онкопротеином Е1А. Nature 396, 184–186
33 Shih, IM, Yu, J., He, TC, Vogelstein, B. and Kinzler, KW (2000) Связывающий домен β-катенина
аденоматозного полипоза толстой кишки достаточен для подавления опухоли .
Рак Рез.60, 1671–1676
34 Arany, Z., Sellers, W.R., Livingston, D.M. and Eckner, R. (1994) E1A-ассоциированные p300 и
CREB-ассоциированные CBP принадлежат к консервативному семейству коактиваторов. Cell 77, 799–800
35 Neish, A.S., Anderson, S.F., Schlegel, B.P., Wei, W. and Parvin, J.D. (1998) Factors
, связанные с холоферментом РНК-полимеразы II млекопитающих. Нуклеиновые Кислоты Res. 26,
847–853
36 Tutter, A. V., Fryer, C.J. and Jones, K.A. (2001) Хроматин-специфическая регуляция
V., Fryer, C.J. and Jones, K.A. (2001) Хроматин-специфическая регуляция
Активация и ингибирование транскрипции LEF-1-β-catenin
in vitro
Гены Дев. 15,3342–3354
37 Townsley, F.M., Cliffe, A. and Bienz, M. (2004) Pygopus and Legless нацеливают
Armadillo/β-catenin на ядро, чтобы обеспечить его транскрипционную функцию коактиватора.
Нац. Клеточная биол. 6, 626–633
38 Mosimann, C., Hausmann, G. and Basler, K. (2006) Parafibromin/Hyrax активирует транскрипцию целевого гена Wnt/Wg
путем прямой ассоциации с β-catenin/Armadillo. Сотовый 125,
327–341
39 Ким, С., Xu, X., Hecht, A. и Boyer, T.G. (2006) Медиатор является преобразователем передачи сигналов
Wnt/β-catenin. Дж. Биол. хим. 281, 14066–14075
Поступила в редакцию 26 января 2009 г./17 июня 2009 г.; Принято 25 июня 2009 г.
Опубликовано как BJ Немедленная публикация 25 июня 2009 г., DOI: 10.1042 / BJ200
C
C
C
C
C
RUB
2009 Биохимическое общество
ACP — Отношения — Аэрозольные оптические свойства, полученные из дракона Кампания -NE Asia и последствия для одноканального алгоритма для получения оптической толщины аэрозоля весной с помощью метеорологического формирователя изображений (MI) на борту спутника связи, океана и метеорологии (COMS)
. Сезонность концентрации частиц и распределения по размерам: глобальный анализ, полученный из сети приповерхностных обсерваторий Глобальной службы атмосферы (ГСА)
Сезонность концентрации частиц и распределения по размерам: глобальный анализ, полученный из сети приповерхностных обсерваторий Глобальной службы атмосферы (ГСА) Клеманс Роуз, Мартина Колло Коэн, Элизабет Эндрюс, Йонг Лин, Исалин Боссер, Катрин Лунд Мире, Томас Туч, Альфред Виденсолер, Маркус Фибиг, Паси Аалто, Андрес Аластуэй, Элизабет Алонсо-Бланко, Маркос Андраде, Бегонья Артинано, Тодор Арсов, Урс Балтеншпергер, Сюзанна Бастиан, Олаф Бат, Йохан Пол Бьюкес, Бенджамин Т.Брем, Николя Буковецки, Хуан Андрес Каскеро-Вера, Себастьен Кониль, Константинос Элефтериадис, Оливье Фавез, Харальд Флентье, Мария И. Джини, Франсиско Хавьер Гомес-Морено, Мартин Гизель-Беер, Анна Ганнет Халлар, Иво Калапов, Никос Каливитис, Энн Каспер-Гибль, Мелита Кейвуд, Чон Ын Ким, Сан-Ву Ким, Адам Кристенссон, Маркку Кулмала, Хейкки Лихавайнен, Ненг-Хуэй Лин, Хассан Лямани, Анджела Маринони, Себастьяо Мартинс Дос Сантос, Ольга Л.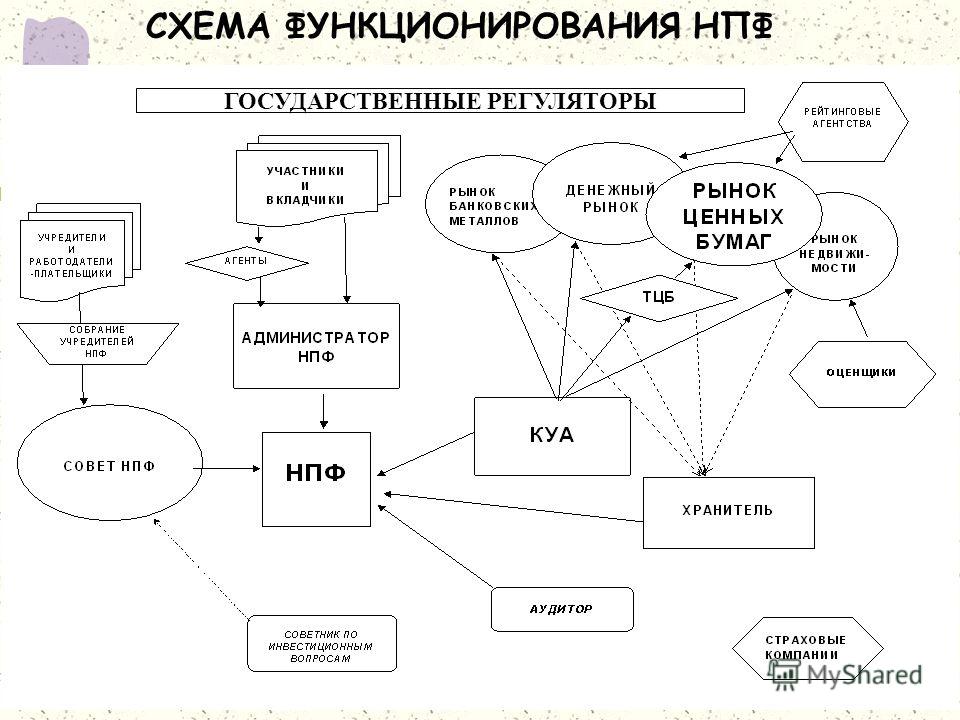 Майол-Брасеро, Фрэнк Мейнхардт , Майк Меркель, Жан-Марк Мецгер, Николаос Михалопулос, Якуб Ондрачек, Марко Пандольфи, Ноэми Перес, Туукка Петая, Жан-Эуд Пети, Давид Пикард, Жан-Марк Пишон, Вероник Пон, Жан-Филипп Путо, Фабьен Райзен, Карин Селлегри , Сангита Шарма, Герхард Шауэр, Патрик Шеридан, Джеймс Патрик Шерман, Андреас Шверин, Ральф Сомер, Мар Соррибас, Джуньинг Сан, Пьер Туле, Вилле Ваккари, Питер Гидеон ван Зил, Фернандо Веларде, Паоло Виллани, Стергиос Вратолис, Зденек Вагнер, Шэн -Сян Ван, Кей Вейнхолд, Рольф Веллер, Маргарита Йела, Владимир Здималь и Паоло Лай
Майол-Брасеро, Фрэнк Мейнхардт , Майк Меркель, Жан-Марк Мецгер, Николаос Михалопулос, Якуб Ондрачек, Марко Пандольфи, Ноэми Перес, Туукка Петая, Жан-Эуд Пети, Давид Пикард, Жан-Марк Пишон, Вероник Пон, Жан-Филипп Путо, Фабьен Райзен, Карин Селлегри , Сангита Шарма, Герхард Шауэр, Патрик Шеридан, Джеймс Патрик Шерман, Андреас Шверин, Ральф Сомер, Мар Соррибас, Джуньинг Сан, Пьер Туле, Вилле Ваккари, Питер Гидеон ван Зил, Фернандо Веларде, Паоло Виллани, Стергиос Вратолис, Зденек Вагнер, Шэн -Сян Ван, Кей Вейнхолд, Рольф Веллер, Маргарита Йела, Владимир Здималь и Паоло Лай
Атмос.хим. Phys., 21, 17185–17223, https://doi.org/10.5194/acp-21-17185-2021, https://doi.org/10.5194/acp-21-17185-2021, 2021
Краткое содержаниеACP — Взаимосвязи — Оптические свойства аэрозолей и прямое радиационное воздействие на основе измерений Китайской сети дистанционного зондирования аэрозолей (CARSNET) в восточном Китае
Введение в специальный выпуск «Углубленное изучение источников и процессов загрязнения воздуха в Пекине и его окрестностях (APHH-Пекин)» Зонгбо Ши, Туан Ву, Симона Коттхаус, Рой М. Harrison, Sue Grimmond, Siyao Yue, Tong Zhu, James Lee, Yiqun Han, Matthias Demuzere, Rachel E. Dunmore, Lujie Ren, Di Liu, Yuanlin Wang, Oliver Wild, James Allan, W. Joe Acton, Janet Barlow, Benjamin Barratt, David Beddows, William J. Bloss, Giulia Calzolai, David Carruthers, David C. Carslaw, Queenie Chan, Lia Chatzidiakou, Yang Chen, Leigh Crilley, Hugh Coe, Tie Dai, Ruth Doherty, Fengkui Duan, Pingqing Fu, Baozhu Ge, Maofa Ge, Daobo Guan, Jacqueline F. Hamilton, Kebin He, Mathew Heal, Dwayne Heard, C.Nicholas Hewitt, Michael Hollaway, Min Hu, Dongsheng Ji, Xujiang Jiang, Rod Jones, Markus Kalberer, Frank J. Kelly, Louisa Kramer, Ben Langford, Chun Lin, Alastair C. Lewis, Jie Li, Weijun Li, Huan Liu, Junfeng Liu, Miranda Loh, Keding Lu, Franco Lucarelli, Graham Mann, Gordon McFiggans, Mark R. Miller, Graham Mills, Paul Monk, Eiko Nemitz, Fionna O’Connor, Bin Ouyang, Paul I. Palmer, Carl Percival, Olalekan Popoola, Claire Reeves, Andrew R. Rickard, Longyi Shao, Guangyu Shi, Dominick Spracklen, David Stevenson, Yele Sun, Zhiwei Sun, Shu Tao, Shengrui Tong, Qingqing Wang, Wenhua Wang, Xinming Wang, Xuejun Wang, Zifang Wang, Lianfang Wei, Lisa Whalley, Xuefang Wu, Zhijun Wu, Pinhua Xie, Fumo Yang, Qiang Zhang, Yanli Zhang, Yuanhang Zhang, and Mei Zheng
Harrison, Sue Grimmond, Siyao Yue, Tong Zhu, James Lee, Yiqun Han, Matthias Demuzere, Rachel E. Dunmore, Lujie Ren, Di Liu, Yuanlin Wang, Oliver Wild, James Allan, W. Joe Acton, Janet Barlow, Benjamin Barratt, David Beddows, William J. Bloss, Giulia Calzolai, David Carruthers, David C. Carslaw, Queenie Chan, Lia Chatzidiakou, Yang Chen, Leigh Crilley, Hugh Coe, Tie Dai, Ruth Doherty, Fengkui Duan, Pingqing Fu, Baozhu Ge, Maofa Ge, Daobo Guan, Jacqueline F. Hamilton, Kebin He, Mathew Heal, Dwayne Heard, C.Nicholas Hewitt, Michael Hollaway, Min Hu, Dongsheng Ji, Xujiang Jiang, Rod Jones, Markus Kalberer, Frank J. Kelly, Louisa Kramer, Ben Langford, Chun Lin, Alastair C. Lewis, Jie Li, Weijun Li, Huan Liu, Junfeng Liu, Miranda Loh, Keding Lu, Franco Lucarelli, Graham Mann, Gordon McFiggans, Mark R. Miller, Graham Mills, Paul Monk, Eiko Nemitz, Fionna O’Connor, Bin Ouyang, Paul I. Palmer, Carl Percival, Olalekan Popoola, Claire Reeves, Andrew R. Rickard, Longyi Shao, Guangyu Shi, Dominick Spracklen, David Stevenson, Yele Sun, Zhiwei Sun, Shu Tao, Shengrui Tong, Qingqing Wang, Wenhua Wang, Xinming Wang, Xuejun Wang, Zifang Wang, Lianfang Wei, Lisa Whalley, Xuefang Wu, Zhijun Wu, Pinhua Xie, Fumo Yang, Qiang Zhang, Yanli Zhang, Yuanhang Zhang, and Mei Zheng
Atmos.